Вредный ген

Задача
Один из этапов получения генно-модифицированных микроорганизмов — отбор (селекция) клеток, успешно встроивших в свой геном или захвативших в качестве самостоятельно реплицирующегося элемента определенный набор генов. Часто процент таких клеток очень низок — всего 1 из 1 000 000; их требуется отобрать из огромного числа других клеток того же штамма, не содержащих этой «генетической конструкции». Для этого обычно используют так называемые «маркерные» гены. Эти гены могут, например, кодировать ферменты, которые разлагают определенный антибиотик. Если поместить смесь клеток в среду, содержащую этот антибиотик, то на ней выживут и дадут потомство только те клетки, в которых есть соответствующий маркерный ген, а остальные — погибнут.
Давайте назовем это «прямой» селекцией: мы отбираем только те клетки, в которых работает определенный ген. Однако иногда бывает необходимость провести и «обратную» селекцию — отобрать те клетки, в которых нет заданного гена или заданный ген не работает. Под словами «ген не работает» мы подразумеваем наличие в этом гене такой мутации, которая не позволяет правильно функционировать закодированному им белку. Обычно маркерные гены позволяют проводить «прямую» селекцию, однако есть и гены, для которых легко проверять их отсутствие. Есть даже гены, позволяющие проводить как «прямую», так и «обратную» селекцию. Такие гены — очень удобный инструмент генной инженерии.
Как вы думаете, каков может быть принцип обратной селекции? Какие функции могут выполнять белки, закодированные маркерными генами для обратной селекции? А что должен делать белок, закодированный геном-маркером, который годится и для прямой, и для обратной селекции?
Подсказка 1
У пекарских дрожжей один из генов, позволяющих проводить «обратную» селекцию, кодирует белок, который встраивается в цитоплазматическую мембрану клетки.
Подсказка 2
Некоторые «маркерные» гены дают возможность клеткам синтезировать аминокислоты или нуклеотиды, которые они не способны синтезировать в отсутствие этого гена.
Решение

Примером гена, позволяющим проводить «обратную» селекцию, является переносчик аминокислоты аргинина, локализованный в цитоплазматической мембране. Рассмотрим этот вопрос подробнее на примере пекарских дрожжей Saccharomyces cerevisiae. Клетки дикого типа дрожжей способны синтезировать аргинин самостоятельно, но также они способны получать аргинин из внешней среды. Для этого они используют переносчик аргинина Can1p, который закодирован геном CAN1. Однако этот же белок способен переносить из внешней среды токсичный для клетки канаванин (см. рис. 1), химическая структура которого крайне похожа на структуру аргинина. Если канаванин попадает в цитоплазму, то он ингибирует собственный синтез аргинина клетками и в конечном счете ее губит. Очевидно, что клетки, в которых отсутствует ген CAN1, не чувствительны к канаванину и спокойно живут в его присутствии. Таким образом, поместив клетки в питательную среду, содержащую канаванин, можно отобрать те, в которых ген CAN1 не работает.
В качестве примера маркерного гена, позволяющего проводить как прямую, так и обратную селекцию, можно рассмотреть ген URA3 дрожжей. Этот ген кодирует фермент, необходимый для синтеза пиримидиновых нуклеотидов, входящих в состав как РНК, так и ДНК — молекул являющихся основой существования каждой живой клетки. Без этого фермента клетки становятся полностью зависимы от наличия в среде экзогенного (добавленного снаружи) урацила. Следовательно, достаточно просто отобрать клетки дрожжей, не содержащие этот ген. Достаточно поместить суспензию клеток на твердую среду, которая содержит все необходимые для роста дрожжей питательные и минеральные вещества кроме урацила. Очевидно, что на такой среде вырастут (размножатся и образуют колонии) только те дрожжи, в которых содержится функциональная копия гена URA3.
Как же устроена обратная селекция? Так же, как и в случае с CAN1 и канаванином, фермент Ura3p (оротодин 5'-фосфат декарбоксилаза) может «ошибаться» и «работать» с молекулой, немного отличающейся от оротидин 5'-фосфата — его привычного субстрата (см. рис. 2). Фермент Ura3p (закодированный геном URA3) способен катализировать реакцию, в которой в качестве субстрата выступает фтороротидин 5'-фосфат, а в качестве одного из продуктов получается крайне токсичный для клетки фторуридин 5'-фосфат и продукт его дальнейшего метаболического преобразования фторурацил (см. рис. 2).
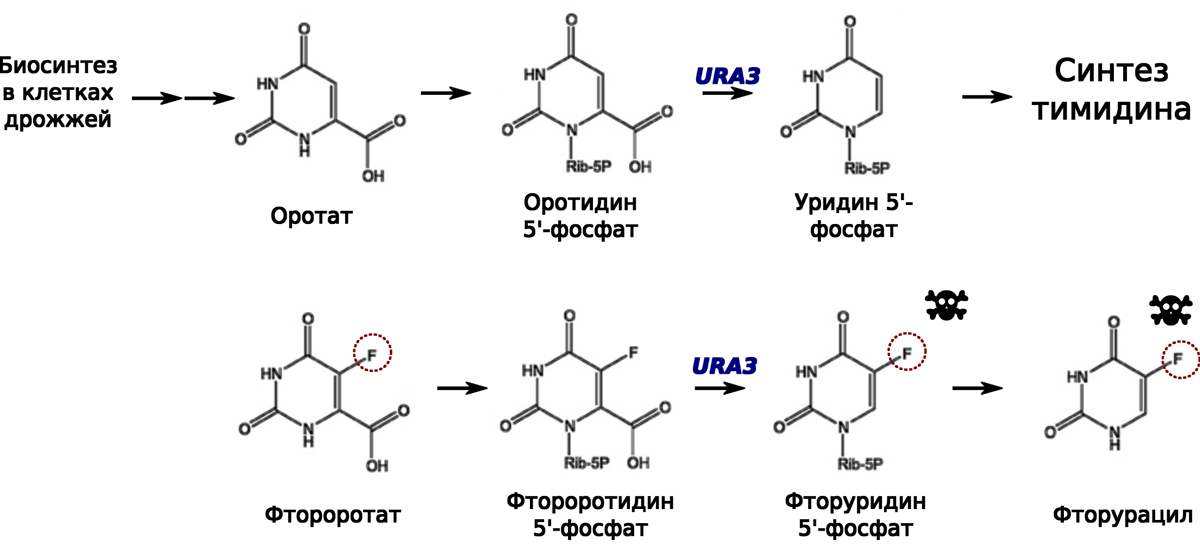
Рис. 2. Метаболические реакции, протекающие при участии URA3. Фтороротат в результате нескольких метаболических реакций
Такое явление — синтез собственными ферментами клетки крайне токсичного соединения — называется «летальный синтез». Предполагается, что фторурацил в клетке полностью блокирует синтез дезокситимидина, полностью блокируя фермент тимидинсинтазу. Поскольку дезокситимидин нужен для синтеза и репарации ДНК, клетки дрожжей, в которых образовывается фторуарацил, не могут нормально жить, а тем более размножаться. Таким образом, если мы добавим к суспензии клеток дрожжей фтороротат, то он не позволит делиться тем клеткам, в которых есть активный фермент Ura3p, а следовательно, есть рабочая копия гена URA3. Это позволяет проводить обратную селекцию, отбирая только те клетки, в которых гена URA3 нет.
Послесловие
В каких случаях возникает необходимость проводить «обратную» селекцию? Обратная селекция позволяет оценивать частоты возникновения мутаций в ядерной ДНК. Если взять клетки дрожжей с работающим геном CAN1 (URA3 также подойдет для подобной задачи) и высадить их на твердую среду, содержащую канаванин (рис. 3), то колонии смогут образовать только те клетки, в которых ген CAN1 не работает. Аналогичный эксперимент можно провести, предварительно проинкубировав клетки с каким-либо химическим соединением. Если это вещество окажется мутагеном, например, будет взаимодействовать с ДНК или мешать работе ферментов репарации, то число колоний на среде с канаванином окажется значительно больше. На рис. 3 приведена схема типичного эксперимента на клетках дрожжей Saccharomyces cerevisiae. Почему возрастет количество колоний в присутствие мутагена? В клетках дрожжей, проинкубированных с мутагеном, с высокой вероятностью возникают изменения нуклеотидной последовательности в разных участках ядерной ДНК. Часть из них «попадает» в ген CAN1. Некоторые из этих изменений могут привести к тому, что белок, закодированный геном CAN1, окажется неспособным выполнять свою функцию — закачивать аргинин и канаванин. Клетка с такой мутацией способна расти и размножаться на среде даже с высоким содержанием канаванина. В результате окажется, что чем выше вероятность повреждения ДНК, тем большее число клеток получит необходимые изменения в гене и образуют колонию на среде с канаванином.

Рис. 3.
Рассмотрим еще один пример. Иногда бывает необходимо исследовать химическое соединение, нарушающее взаимодействие двух белков. Такой поиск обычно является одним из начальных этапов создания новых лекарств. Пусть, например, нам надо найти химическое соединение, нарушающее связывание белков X и Y. Для этого нужно сконструировать два гена, кодирующих два химерных белка (рис. 4), каждый из которых составлен из двух частей (доменов). Первый белок должен состоять из домена, связывающегося с определенной последовательностью ДНК (GAL DBD — от Galactose DNA Binding Domain) и белка X. Второй белок — из белка Y и специального домена, необходимого для нормальной работы маркерного гена (GAL AD — от Galactose Activator Domain). В качестве такого гена как раз и пригодится маркер, позволяющий проводить обратную селекцию. В этой системе, ген будет работать только в том случае, если белки X и Y будут прочно связываться друг с другом и «привлекать» белок GAL AD к необходимому участку ДНК в регуляторной области гена URA3.
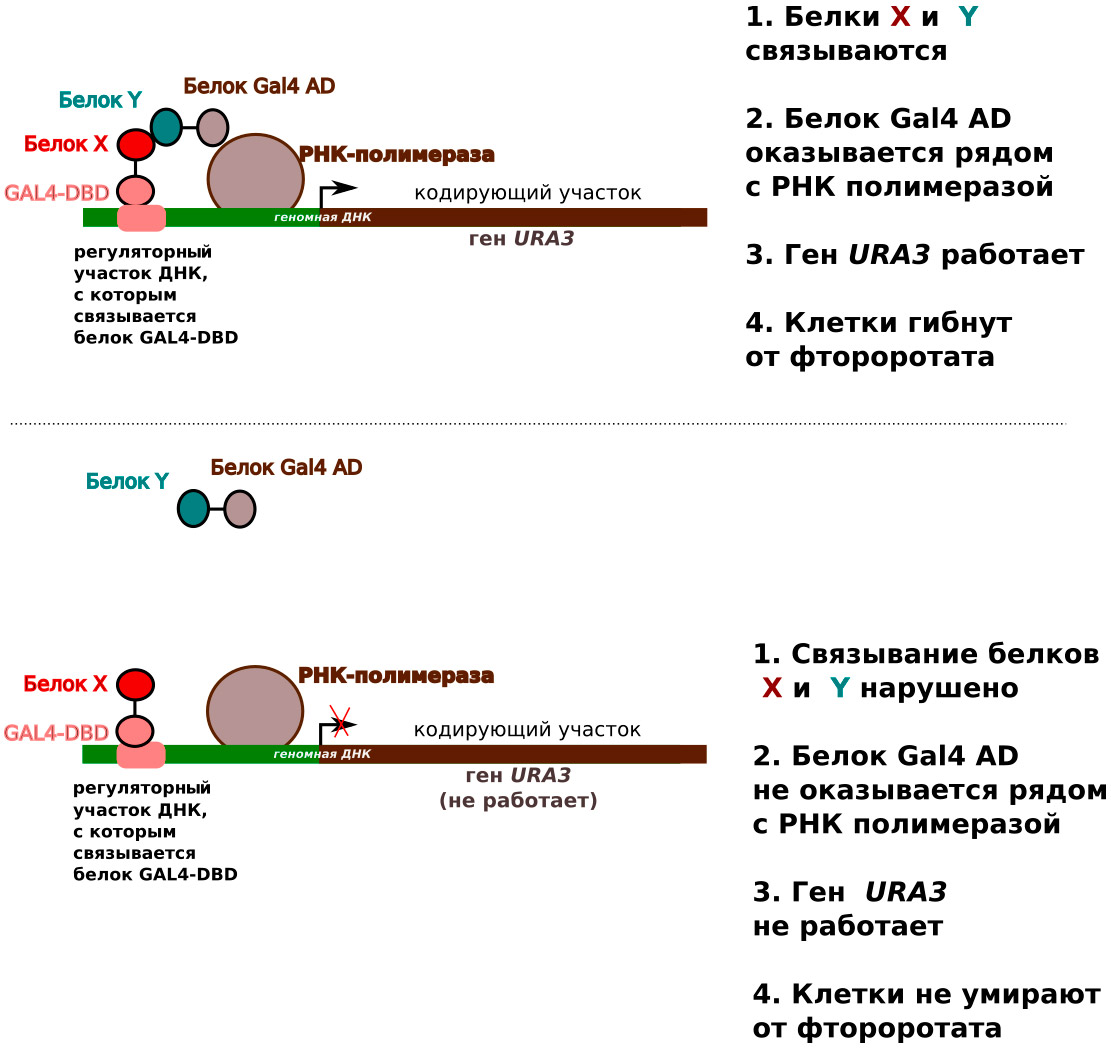
Рис. 4.





Рис. 1. Химическая структура канаванина (сверху) и аргинина (снизу)