Найден маленький рибозим-полимераза, способный реплицировать сам себя
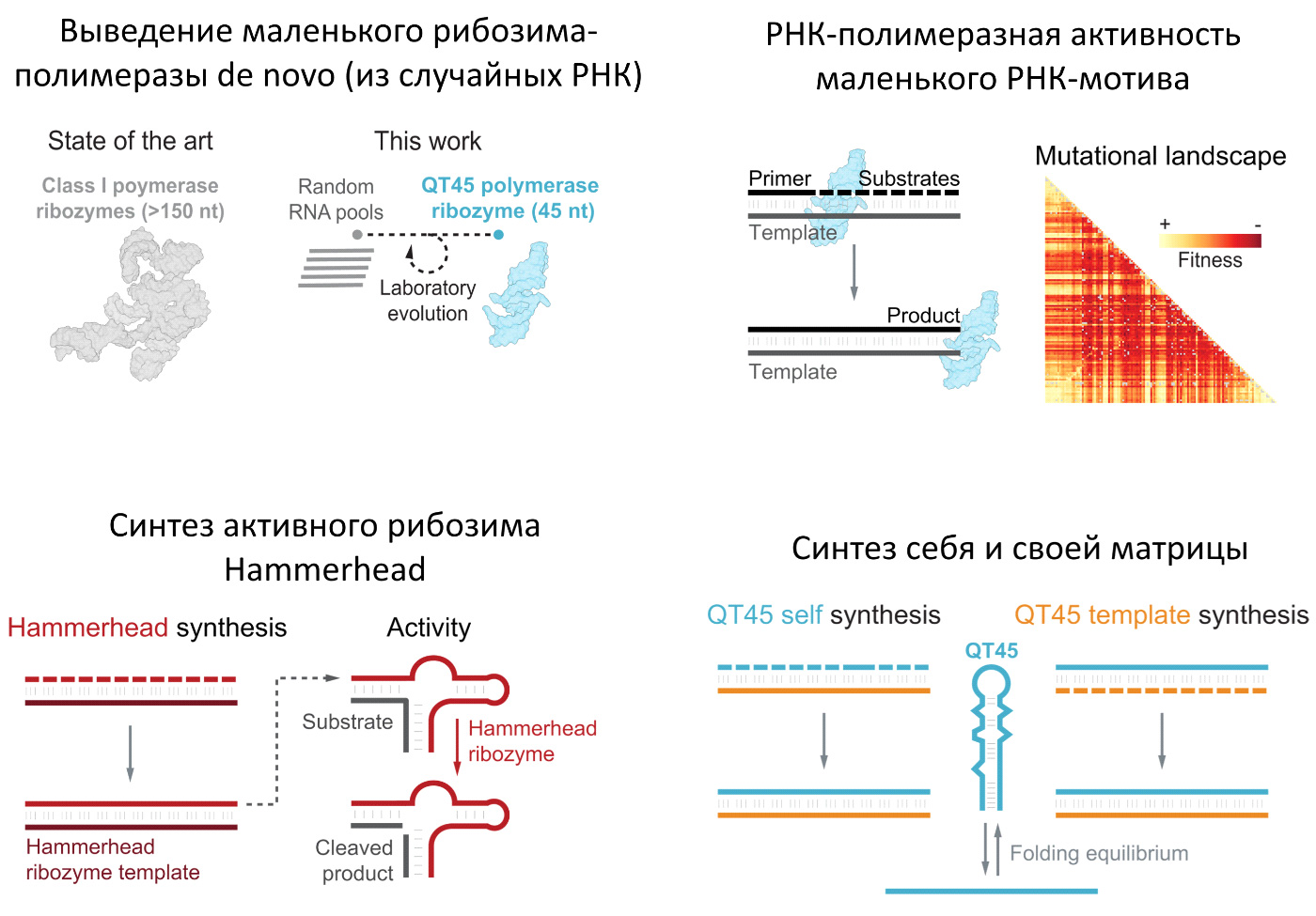
Согласно теории РНК-мира, на ранних этапах абиогенеза важную роль играли рибозимы-полимеразы — молекулы РНК, способные размножать молекулы РНК, в том числе самих себя. До сих пор ученым удавалось получить (вывести путем искусственной эволюции или целенаправленно спроектировать) только крупные рибозимы-полимеразы длиной более 150 нуклеотидов, ни один из которых не мог реплицировать сам себя. Вероятность случайной самосборки таких больших молекул исчезающе мала, сами они недолговечны, а для их устойчивого размножения и дальнейшей эволюции необходима труднодостижимая скорость и точность репликации. Британские ученые сделали важный шаг к преодолению этих трудностей. Использовав новые методы отбора, они вывели из случайных молекул РНК маленький рибозим-полимеразу QT45 длиной всего в 45 нуклеотидов, способный к саморепликации. Исследование показало, что в пространстве последовательностей РНК мотивы с полимеразной активностью встречаются намного чаще, чем предполагалось. А главное, среди них есть совсем маленькие. Это делает более правдоподобной идею об их эволюционном возникновении посредством неферментативной репликации и даже позволяет допустить случайную самосборку готового рибозима-полимеразы.
1. Роль рибозимов-полимераз в мире РНК
Последние достижения в развитии теории мира РНК (см. ссылки в конце новости) позволяют считать, что начальные этапы зарождения жизни, вероятнее всего, были следующими:
1) Абиогенный синтез нуклеотидов в геотермальных системах из смеси простейших абиогенных углеводов и азотистых соединений.
2) Олигомеризация: синтез случайных коротких олигонуклеотидов на минеральных матрицах, с возможным участием аминокислот в качестве катализаторов и важной ролью циклов обводнения-высыхания в геотермальных системах.
3) Неферментативная репликация — возможно, путем матричного лигирования в слабощелочных условиях (см. Аминокислоты помогают нуклеотидам соединяться в короткие молекулы РНК, «Элементы», 16.07.2025). Старт дарвиновской эволюции.
4) Эволюция рибозимов, в том числе полимераз — молекул РНК, катализирующих размножение (репликацию) других молекул РНК и самих себя. Рибозимы-полимеразы, эволюционируя, повышают скорость и точность репликации, что подстегивает прогрессивную эволюцию живых систем.
5) Эволюция сложных РНК-организмов и программируемого синтеза белков.
Если бы этап 2 протекал очень бурно, а первые эффективные рибозимы из этапа 4 были совсем маленькими, то в принципе можно было бы даже пропустить этап 3. Иначе говоря, первые достаточно эффективные рибозимы-полимеразы могли бы собраться из нуклеотидов «случайно», без помощи наследственности и отбора.
В любом случае рибозимы-полимеразы должны были играть ключевую роль на определенном этапе абиогенеза. Поэтому их активно изучают. У современных живых организмов таких рибозимов нет. Их давно вытеснили гораздо более эффективные белковые полимеразы. Но в прошлом они были, а значит, их можно воссоздать.
Ученые ищут последовательности нуклеотидов с полимеразной активностью в бескрайнем пространстве последовательностей (см. Sequence space (evolution)), выбирая перспективных кандидатов из множества случайных молекул РНК. Затем пытаются улучшить рабочие качества этих кандидатов при помощи искусственной эволюции (внесение случайных мутаций — отбор наилучших — следующий раунд размножения с мутациями и т. д.), комбинируя эволюционный подход с элементами сознательного проектирования (см. Рибозимы могут размножать друг друга, «Элементы», 13.04.2011).
Год за годом мы наблюдаем в этой области впечатляющий прогресс. Например, недавно удалось повысить точность работы рибозима-полимеразы до уровня, достаточного для обеспечения настоящей прогрессивной эволюции другого рибозима, правда очень маленького — эндонуклеазы Hammerhead (см. Эволюция рибозимов, размножаемых рибозимами: еще один шаг к воссозданию РНК-жизни в пробирке, «Элементы», 27.03.2024).
2. Почему так важно найти маленький рибозим-полимеразу
Все достижения в изучении рибозимов-полимераз до сих пор были связаны с большими, громоздкими молекулами длиной более 150 нуклеотидов. Начинало даже закрадываться подозрение, что молекула РНК меньшего размера вообще не может иметь полимеразной активности. Это вело к пессимистичным (то есть низким) оценкам вероятности абиогенеза и частоты встречаемости жизни во Вселенной по нескольким причинам:
1) Вероятность случайной самосборки конкретной последовательности из 150 нуклеотидов исчезающе мала. Возможных последовательностей такой длины существует 4150 ≈ 1090. Это сопоставимо с числом элементарных частиц в наблюдаемой Вселенной. Дарвиновская эволюция на основе неферментативной репликации тоже вряд ли смогла бы вывести такого 150-нуклеотидного монстра, если до достижения этого размера он не помогал реплицировать РНК.
2) Чтобы 150-нуклеотидная молекула могла нормально воспроизводиться и эволюционировать, не деградируя и накапливая полезные мутации, требуется очень высокая точность репликации. Нужно, чтобы в среднем было не более двух-трех ошибок на копию рибозима. Рибозимы-полимеразы, способные синтезировать 150-нуклеотидную молекулу с такой точностью, пока не найдены.
3) Даже без учета требований к точности репликации, рибозимам-полимеразам трудно реплицировать длинные матрицы, особенно если в них используются все 4 нуклеотида в произвольном порядке. Поэтому ни один из известных рибозимов-полимераз до сих пор не мог реплицировать сам себя (хотя некоторые подошли к этому довольно близко окольными путями, см. K. F. Tjhung et al., 2020. An RNA polymerase ribozyme that synthesizes its own ancestor).
4) Молекулы РНК недолговечны, в том числе в условиях, считающихся «пребиотически правдоподобными». Длинные молекулы, как правило, деградируют быстрее коротких. Это повышает требования к скорости работы полимеразы. Ведь для устойчивого самовоспроизводства она должна успеть за свою недолгую жизнь создать хотя бы одну-две полноценные копии самой себя. Известные рибозимы-полимеразы неторопливы: для копирования одной длинной молекулы РНК им обычно требуются даже не часы, а дни, что сопоставимо с периодом полураспада самого рибозима.
В свете этих фактов понятно желание исследователей под руководством Филиппа Холлигера (Philipp Holliger), известного специалиста по абиогенезу, найти радикально более короткий рибозим-полимеразу. Если земная жизнь — не нечто абсолютно уникальное, что может случайно появиться лишь в одной из бесчисленных вселенных Мультиверса, в пространстве последовательностей РНК должны существовать более короткие и изящные полимеразы, чем те неулюжие 150–200-нуклеотидные молекулы, с которыми ученые возились до сих пор.
3. Как родился совсем крохотный рибозим QT45
Исследователи не стали пытаться укоротить какую-то из известных полимераз. Они решились на более смелый шаг и начали поиск «с нуля», то есть с набора случайных коротких последовательностей. Точнее, с трех наборов случайных РНК разной длины: по 20, 30 и 40 нуклеотидов. В каждом наборе было примерно по шесть триллионов (6 · 1012) уникальных последовательностей. Это соответствует полному покрытию всего пространства последовательностей только для самых коротких молекул длиной в 20 нуклеотидов. Для 30-нуклеотидных молекул было охвачено лишь несколько миллионных, для 40-нуклеотидных — несколько триллионных от общего числа возможных последовательностей.
Первичный отбор последовательностей, обладающих хотя бы намеком на полимеразную активность, производился следующим образом (рис. 2). К каждой случайной молекуле (потенциальному рибозиму) присоединяли через гибкий РНК-овый мостик молекулу «субстрата» (олигонуклеотид, активированный тремя фосфатами на 5’-конце). Добавляли праймер (другой олигонуклеотид с молекулой биотина на 5’-конце) и матрицу — молекулу РНК, комплементарную субстрату и праймеру. Если потенциальный рибозим успешно проводил лигирование, то есть пришивал субстрат (а заодно и самого себя) к праймеру, то этот рибозим потом отлавливали благодаря наличию биотина на 5’-конце праймера. Это классическая методика, основанная на том, что биотин прилипает к шарикам, покрытым биотин-связывающим белком стрептавидином.

Рис. 2. Конструкция, использовавшаяся для первичного отбора последовательностей с минимальной способностью сшивать (лигировать) олигонуклеотиды, комплементарные матрице. Рисунок из обсуждаемой статьи.
Перед отобранными таким способом последовательностями в последующих раундах отбора ставили всё более сложные задачи, постепенно приближая отбираемые рибозимы к конечной цели: способности реплицировать любые матрицы путем последовательного присоединения комплементарных тринуклеотидов (триплетов).
Исследователи использовали в качестве «субстратов» не отдельные нуклеотиды, а триплеты, потому что из множества предыдущих работ постепенно становится ясно, что это — перспективный путь. Вот некоторые соображения в пользу триплетов (или других коротких олигомеров, но не одиночных нуклеотидов) в качестве субстратов для репликации в РНК-мире:
1) Триплеты комплементарно присоединяются к матрице в разных местах, мешая матрице сворачиваться в шпильки. Это упрощает репликацию молекул РНК со сложной трехмерной структурой.
2) Тот же механизм помогает справляться с проблемой повторного спаривания матриц с синтезированными на них комплементарными репликами.
3) Разных триплетов не так много — всего 64. Гипотеза о том, что все они могли в достаточной концентрации присутствовать в «колыбели жизни», не выглядит невероятной (см. Аминокислоты помогают нуклеотидам соединяться в короткие молекулы РНК, «Элементы», 16.07.2025).
4) Если эволюция репликации РНК начиналась с неферментативного матричного лигирования олигонуклеотидов, то логично, что первые рибозимы — РНК-полимеразы — тоже продолжали работать с олигонуклеотидами. Им незачем было переключаться на одиночные нуклеотиды, с которыми у них сразу стало бы больше проблем.
5) Есть интереснейшая линия гипотетических построений, согласно которой в РНК-мире существовала трипликаза — рибозим, который реплицировал РНК путем последовательного присоединения триплетов. Возможно, трипликаза отрезала эти триплеты от специальных маленьких РНК — переносчиков триплетов и по совместительству предков тРНК. Этот механизм репликации впоследствии лег в основу механизма трансляции, а сама трипликаза стала малой субъединицей рибосомы (см. D. Penny, 2005. An interpretive review of the origin of life research; критический обзор в: Y. I. Wolf & E. V. Koonin, 2007. On the origin of the translation system and the genetic code in the RNA world by means of natural selection, exaptation, and subfunctionalization). Гипотеза трипликазы родилась из изучения рибосомы и попыток понять происхождение трансляции. Изначально она никак не была связана с тем, что специалисты по неферментативной репликации и рибозимам-полимеразам заметили, что с триплетами работать проще, чем с одиночными нуклеотидами.
За 11 раундов отбора исследователям удалось вывести три непохожих друг на друга коротких рибозима-полимеразы, худо-бедно справляющихся с репликацией разнообразных матриц. Все три происходили из набора 40-нуклеотидных последовательностей. 20- и 30-нулеотидные молекулы, по-видимому, всё-таки слишком коротки, чтобы проявлять полимеразную активность.
С этими тремя кандидатами стали работать дальше, внося случайные мутации, ставя всё более жесткие критерии отбора и добавляя капельку сознательного проектирования. После семи дополнительных раундов отбора (суммарно — 18 раундов) исследователи пришли к симпатичному 51-нуклеотидному рибозиму-полимеразе (рис. 3, B), который потом удалось сократить до 45-нуклеотидного почти без ущерба для функциональности. Итоговый рибозим получил название QT45 (QT — аббревиатура слов Quite Tiny — «cовсем крохотный»).

Рис. 3. Сравнение одного из лучших полученных ранее рибозимов-полимераз 5TU/t1.5 (димер из двух молекул РНК длиной 152 и 136 нуклеотидов) и нового 51-нуклеотидного рибозима QT51 (который потом сократили до 45-нуклеотидного QT45). Рисунок из дополнительных материалов к обсуждаемой статье
Успеху способствовали следующие удачные решения:
- Авторы не пытались «довести до ума» и сократить уже известные огромные рибозимы-полимеразы, а провели целенаправленный поиск коротких мотивов буквально с нуля, из хаоса, как в первый день Творения. Это позволило довольно тщательно прочесать пространство коротких нуклеотидных последовательностей.
- Использование условий, облегчающих работу рибозимов-полимераз: реакции проходили в слабощелочном эвтектическом льду (см. Eutectic system). В заполненных жидкостью микрополостях льда достигается высокая концентрация реагентов, а рибозимы на холоде более стабильны.
- Выбор в качестве субстратов не мононуклеотидов, а триплетов (см. выше).
- Удачная схема отбора с гибким соединительным мостиком и биотинилированным праймером (рис. 2). Это позволило превратить даже слабую каталитическую активность в надежно отлавливаемый сигнал.
4. Свойства рибозима QT45
QT45 — представитель совершенно нового класса рибозимов-полимераз, не похожий по своей структуре на изучавшиеся ранее. Он может реплицировать матрицы с практически любой последовательностью нуклеотидов, потому что его работа не связана с образованием каких-либо Уотсон-Криковских связей (см. Спаренные основания) с комплексом «матрица-праймер-субстрат».
Дополнительные эксперименты показали, что для правильного взаимодействия QT45 с этим комплексом важны 2’-OH группы (то есть гидроксильные группы, присоединенные ко второму атому углерода в рибозе, см. Структра РНК) некоторых нуклеотидов матрицы, праймера и субстрата, расположенных на расстоянии от 1 до 4 нуклеотидных позиций от места лигирования. Похожая картина характерна и для некоторых больших рибозимов-полимераз, изучавшихся ранее. По-видимому, это говорит о том, что разные классы рибозимов-полимераз имеют конвергентно сходные способы обхватывания комплекса «матрица-праймер-субстрат» и взаимодействия с ним. Также из этого следует, что эти рибозимы вряд ли подходят для репликации ДНК, ведь наличие 2’-OH групп — это как раз то, что отличает рибонуклеотиды от дезоксирибонуклеотидов.
QT45 не слишком привередлив в выборе субстратов. Он может использовать для синтеза комплементарной нити РНК не только триплеты, но и более длинные олигонуклеотиды, активированные тремя фосфатами на 5’-конце. Также он может, хоть и с меньшим успехом, использовать динуклеотиды и даже отдельные мононуклеотид-трифосфаты. Не возражает он и против смесей всех этих субстратов в разных пропорциях, а также триплетов, активированных не тремя фосфатами, а двумя фосфатами и аденозином (что считается чуть более «пребиотически правдоподобным» вариантом). Такая неразборчивость делает более весомым предположение, что рибозимы, подобные QT45, могли участвовать в ранних этапах эволюции РНК-мира в «пребиотическом бульоне», где было намешано много всякой абиогенной органики.
Исследователи тщательно изучили ландшафт приспособленности (см. Fitness landscape) QT45. Иными словами, они проверили, как влияют на его «приспособленность» (функциональность) различные мутации. Рибозимы-мутанты должны были реплицировать матрицу, последовательно присоединив к праймеру три триплета UGC. Успешность выполнения этой задачи считалась мерой приспособленности. Объем работы, проделанный исследователями, впечатляет: они проверили все возможные однонуклеотидные замены (3 · 45 = 135 вариантов) и делеции (45 вариантов), а также 98% двойных замен (которых всего возможно 3 · 45 · 3 · 44 = 17 820 вариантов).
Анализ показал, что рибозим QT45 находится на вершине четко выраженного «пика приспособленности». Иными словами, большинство мутаций сильно снижают его функциональность (рис. 4). Но всё-таки не все: например, двойные мутации в участках, образующих двойную спираль, часто оказываются невредными, если сохраняется Уотсон-Криковская связь (например, комплементарная пара A-U заменяется на G-C).

Рис. 4. Влияние одиночных мутаций (однонуклеотидных замен и делеций) на функциональность рибозима-полимеразы QT45. Темно-красный цвет соответствует резкому снижению функциональности, светло-желтый — ее сохранению. Рисунок из обсуждаемой статьи
5. Схлопывание вселенной невероятности (лирическое отступление)
Вероятность случайной самосборки какой-то конкретной 150-нуклеотидной последовательности сопоставима с единицей, деленной на число элементарных частиц во Вселенной, включая фотоны и нейтрино (1090). Попросту говоря, такое событие невероятно, если не привлекать концепцию Мультивселенной, а это нежелательный прием в научном рассуждении. Конечно, в пространстве 150-нуклеотидных последовательностей много разных РНК-полимераз, и это наверняка избавляет нас от десятка-другого порядков невероятности. Но даже 1070 — число довольно безнадежное. Это число нуклонов в крупной галактике. Всё равно похоже на полную невероятность.
Что меняется в этих не очень серьезных (игнорирующих возможность эволюции на основе неферментативной репликации) расчетах благодаря открытию 45-нуклеотидной полимеразы? На первый взгляд может показаться, что разница не принципиальная: подумаешь, размер полимеразы уменьшили втрое. Но это обманчивое впечатление, ведь мы имеем дело с комбинаторикой и степенной функцией. Число всех возможных 45-нуклеотидных последовательностей — 1027. Это не в три, а в 1063 раз меньше, чем число возможных 150-нуклеотидных последовательностей. Это сразу перебрасывает нас из Макрокосма в Микрокосм. 1027 — это уже не про число элементарных частиц во Вселенной, а, скорее, про число атомов в одном-единственном человеческом теле.
С учетом результатов анализа ландшафта приспособленности QT45, его толерантности к мутациям, а также того, что среди примерно 1013 случайных 40-нуклеотидных последовательностей (из 1024 возможных) нашелся даже не один, а целых три разных РНК-полимеразных мотива (два других остались пока неизученными), — с учетом всего этого авторы считают, что можно смело отбросить еще десяток порядков, и тогда вероятность того, что случайная последовательность длиной в 40 или более нуклеотидов будет проявлять заметную РНК-полимеразную активность, можно считать равной примерно 10−16.
Это уже сущие пустяки. 1016 случайных молекул РНК могли содержаться в одном ведре «пребиотического бульона» в какой-нибудь геотермальной системе, где шли процессы, подобные описанным в новости Аминокислоты помогают нуклеотидам соединяться в короткие молекулы РНК («Элементы», 16.07.2025).
«Вселенная невероятности» схлопнулась до размеров геотермальной лужи. Скромной лужи где-нибудь на холодном протоматерике. Почему на холодном? Потому что нам пригодится немного льда по берегам.
6. Первые подвиги QT45: репликация РНК с двуспиральными участками и размножение маленького рибозима
Серьезную трудность для рибозимов-полимераз представляют матрицы с двуспиральными участками. Если эти участки уже скручены в двойную спираль, как к ним будут пристраиваться комплементарные нуклеотиды или триплеты?
С этой трудностью уже умеют справляться некоторые большие рибозимы-полимеразы, выведенные ранее. Справился с ней и крошка QT45. Сначала ему давали произвольные матрицы с двуспиральными участками длиной в 4, 6 и 8 пар нуклеотидов. QT45 успешно реплицировал их, причем выход готового продукта (полных реплик предложенной матрицы) повышался при высоких концентрациях субстратов (триплетов). Дело в том, что для молекул РНК, способных скручиваться в трехмерные структуры с двойными спиралями, обычно характерно некоторое равновесие между скрученным и расправленным состояниями. Иначе говоря, они не всегда остаются скрученными, а иногда немножко раскручиваются. Вот тут-то и вступают в игру триплеты: они комплементарно пристраиваются к подходящим участкам раскрутившейся матрицы и мешают ей скрутиться обратно. Это помогает рибозиму-трипликазе делать свою работу.
Следующим подвигом QT45 стал синтез рабочей копии маленького, 33-нуклеотидного рибозима-эндонуклеазы Hammerhead. Репликация Hammerhead уже стала классикой в подобных исследованиях. Большой рибозим, способный реплицировать Hammerhead, впервые был выведен в 2011 году (см. Рибозимы могут размножать друг друга, «Элементы», 13.04.2011). Ну а сегодня полимеразу, не способную реплицировать Hammerhead, просто не будут воспринимать всерьез. Дошло до того, что некоторые большие рибозимы-полимеразы научились реплицировать Hammerhead с точностью, достаточной для полноценной эволюции без помощи белковых ферментов (см. Эволюция рибозимов, размножаемых рибозимами: еще один шаг к воссозданию РНК-жизни в пробирке, «Элементы», 27.03.2024).
Но большие рибозимы-полимеразы — гиганты по сравнению с Hammerhead. Этого нельзя сказать о QT45, который по размеру и структурной сложности лишь едва превосходит Hammerhead. И всё же QT45 справился с задачей. Правда, трудился он долго: целых 65 дней молчаливой работы в толще льда понадобилось ему, чтобы полностью (то есть на полную длину) отреплицировать матрицы Hammerhead с выходом конечного продукта 1,88%. Это значит, что 1,88% предоставленных ему праймеров были успешного достроены до полноразмерного Hammerhead. Да и это только в случае, когда в качестве субстратов ему дали лишь те 11 триплетов, которые необходимы для данной конкретной задачи. При наличии всех 64 триплетов выход конечного продукта снизился до 0,39%. Зато точность репликации оказалась на высоте: 92,6%. Это даже лучше, чем у многих больших полимераз. Для 33-нуклеотидного рибозима такая точность соответствует среднему числу ошибок 2,44 на одну копию Hammerhead. При этом 8,7% (1/e2,44) копий получаются идеальными — без единой ошибки.
Высокая точность объясняется эффектом «торможения на ошибке»: после присоединения не совсем комплементарного триплета дальнейшая репликация тормозится. В результате средняя точность репликации повышается ценой снижения выхода итогового продукта.
7. Триумф QT45: саморепликация
Размножение самого себя — весьма специфическая задача для рибозима-полимеразы. В некоторых отношениях она более сложна, чем репликация любой другой молекулы РНК той же длины. Отчасти поэтому ни одному рибозиму-полимеразе до сих пор не удавалось справиться с этой задачей. Хотя главная причина, конечно, в огромном размере этих рибозимов. Крохотный размер QT45 дал ему шанс стать первым рибозимом, осуществившим полный цикл саморепликации. И он этот шанс не упустил.
Саморепликация состоит из двух этапов. Сначала рибозим должен синтезировать свою комплементарную реплику (минус-цепь), используя себя в качестве матрицы. Затем он должен на матрице минус-цепи синтезировать копию себя (плюс-цепь).
Синтез минус-цепи ставит перед рибозимом два противоположных требования. С одной стороны, он должен быть скручен в устойчивую трехмерную конфигурацию, чтобы хорошо справляться с функцией рибозима. С другой стороны, он должен быть раскручен, чтобы хорошо справляться с функцией матрицы.
К счастью, молекул рибозима в каждой микрополости эвтектического льда много, и в каждый момент времени одни молекулы находятся в свернутом состоянии, а другие в развернутом. Баланс свернутости-развернутости зависит от концентрации триплетов, потому что триплеты комплементарно пристраиваются к подходящим участкам рибозима и мешают ему сворачиваться. Слишком высокие концентрации триплетов подавляют работу QT45, потому что он не может правильно свернуться, слишком низкие — тоже замедляют ее, потому что рибозиму не хватает субстратов для синтеза минус-цепи. Методом проб и ошибок исследователи подобрали оптимальную концентрацию триплетов и добились того, что рибозим за 72 дня синтезировал полноразмерные минус-цепи с выходом конечного продукта 0,24%. Из полученных минус-цепей 10,9% не содержали ни одной мутации, то есть репликация была довольно точной.
Полдела было сделано.
У второго этапа саморепликации — синтеза плюс-цепи на матрице минус-цепи — есть свой подводный камень. Он заключается в том, что рибозим (плюс-цепь) и матрица (минус-цепь) норовят сплестись в двойную спираль, которая является очень выгодным термодинамически и поэтому устойчивым, но бесполезным тупиковым продуктом.
Игры с концентрациями триплетов не помогли решить эту проблему. Оказалось, что плюс-цепь уверенно побеждает триплеты в конкуренции за право сплестись с минус-цепью. Тогда ученые решили, что нужно добавить в систему, помимо триплетов, такой субстрат, который будет сплетаться с минус-цепью с большим энтузиазмом, чем это делают триплеты. Они попробовали использовать гексамеры — маленькие РНК из шести нуклеотидов, то есть, по сути, два заранее сшитых триплета. Это сработало. Оказалось, что достаточно добавить в исходную смесь один-единственный активированный гексамер (pppAUUGAU, где ppp — три фосфата на 5’-конце), чтобы получить желаемый продукт — полноразмерные плюс-цепи1. Выход готового продукта за 72 дня — 0,17%, доля идеальных плюс-цепей без единой мутации 43,4%.
Таким образом, исследователи показали, что QT45 способен худо-бедно осуществлять оба этапа саморепликации. Он стал первым рибозимом, которому это удалось, а первопроходцам всегда трудно. Поэтому не нужно судить его слишком строго за медлительность и неточность, а лучше поздравить с великим достижением.
8. Что дальше?
Речь пока не идет о повторяющихся полноценных циклах саморепликации. Иными словами, ученые еще не могут завести у себя в лаборатории популяцию рибозимов, которая будет без постоянного вмешательства человека жить, размножаться и при этом не вырождаться под грузом мутаций. Однако путь к достижению этой амбициозной цели уже намечен. Основных проблем две.
Во-первых, чтобы процесс мог продолжаться, нужно своевременно расплетать двойные спирали, получающиеся в результате синтеза плюс-цепи на матрице минус-цепи и наоборот. Для этого нужны циклические изменения температуры и pH: в теплой слабокислой среде расплетаются цепи, в слабощелочном льду идет репликация. Можно представить себе такие циклы в какой-нибудь доисторической геотермальной системе, и, конечно, можно смоделировать их в лаборатории. Группа Холлигера уже научилась это делать, что позволило добиться экспоненциального размножения коротких РНК, реплицируемых большим рибозимом-полимеразой (J. Attwater et al., 2025. Trinucleotide substrates under pH–freeze–thaw cycles enable open-ended exponential RNA replication by a polymerase ribozyme).
Во-вторых, QT45 всё-таки недостаточно точен, чтобы избежать вырождения (мутационного коллапса) уже в первых нескольких циклах саморепликации. Но не будем забывать, что он совсем недавно родился «из ничего», из хаоса случайных последовательностей, от которого его отделяют лишь 18 раундов отбора. Так что с большой вероятностью потенциал рибозима QT45 еще не исчерпан, и его можно существенно улучшить. Пожелаем ему удачи.
Источник: Edoardo Gianni, Samantha L. Y. Kwok, Christopher J. K. Wan, Kevin Goeij, Bryce E. Clifton, Enrico S. Colizzi, James Attwater, Philipp Holliger. A small polymerase ribozyme that can synthesize itself and its complementary strand // Science. 2026. V. 391 (6789). P. 1022–1028.
См. также:
1) Рибозимы могут размножать друг друга, «Элементы», 13.04.2011.
2) Созданы рибозимы, синтезирующие зеркальные копии самих себя, «Элементы», 03.11.2014.
3) Александр Марков. В поисках начала эволюции // «Природа» №1, 2015.
4) Эволюция рибозимов, размножаемых рибозимами: еще один шаг к воссозданию РНК-жизни в пробирке, «Элементы», 27.03.2024.
5) Аминокислоты помогают нуклеотидам соединяться в короткие молекулы РНК, «Элементы», 16.07.2025.
1 Две технические подробности для интересующихся: 1) гексамер AUUGAU — это фрагмент из середины рибозима QT45, на рис. 4 он находится сверху; 2) плюс-цепь, являющуюся продуктом реакции, можно отличить от плюс-цепи, являющейся исходным рибозимом, по приделанному меченому праймеру.
-
Всё же полагаю, что для самозарождения жизни нужны не просто геотермальные источники, а геотермальные источники в приливной зоне. Там к накладывающимся (геотермальный + приливной) циклам обводнения-высыхания добавляются циклы смены химического состава воды. А на ранней Земле зон с таким сочетанием было очень намного больше, чем сейчас т.к. и геотермальных источников было больше и Луна ближе.
-
Прилив тут же размоет всё что набулькал вулкан. Пока рабочая идея, что реакция вообще идёт, только пока рассол пересыхает и обновляется на уровне даже не капель воды. Причём рядом ещё нужен сульфат цинка, ультрафиолет и т.п. Большой объём это всё моментально разнесёт.
-
Размоет прилив или нет, и что именно - это ещё бабушка надвое сказала. Длинные полимеры, насколько я понимаю, обычно не плавают в бульоне, а сидят на субстрате, и их не так просто оторвать. А что прилив заменит "бульон" с плавающей в нём мелочью, так оно, может, и к лучшему: уберет продукты реакции, снова выведя систему из равновесия. При этом в заключительном параграфе уже Александр Марков пишет что "нужны циклические изменения температуры и pH". Периодическое разбавление геотермальной воды океанической как раз циклические изменения pH и может обеспечить, в том числе переходы от щелочной среды к кислотной и обратно. В отличие, кстати, от циклов обводнения-высыхания в геотермальном источнике, при которых перехода через нейтральную точку не происходит.
-
Можно не сомневаться, что размоет. Там не то, что прилива быть не должно, даже просто текущей воды не было. Доказательством тому, что прилив не мог участвовать является хотя бы состав внутриклеточных солей.
субстрате, и их не так просто оторвать
Это ещё бабка надвое сказала, зато всё остальное оторвать и разнести на космические расстояния.уберет продукты реакции, снова выведя систему из равновесия
Ток свободной воды эту систему уничтожит, а не выведет из равновесия.
Там должны быть какие-то капиллярные потоки, безусловно, плюс ещё какие-то реакции генерирующие органику в окружении, но это точно не массив воды."нужны циклические изменения температуры и pH"
Циклические изменения температуры и концентрации на поверхности это естественный процесс. Про pH - мы тут говорим про конкретную исследуемую реакцию, которая врядли была связана с той, которая дала начало жизни. Т.е. не надо думать, что эта работа имеет реальное отношение к происхождению жизни, она скорее некоторые ориентиры общего характера подтверждает.-
Согласен вода была врагом абиогенеза, она разрушала и размывала, а другом был ультрафиолет который уничтожал все что не устойчивое и нереплицируемое. При этом матрица аппатита имеет как ромбические поры где могут укладываться цепочки, так и поверхность с тетраэдрами РО4 которые могут работать как минералосвязанная АТФ. Я честно не понимаю почему упрямо пытаются спуститься от молекул вниз вместо того чтобы идти от минералов вверх. Возможно пугают большие объемы исследований и отсутствие смежных знаний по геохимии.
-
Кажется, что это на порядки сложнее анализировать. Одну молекулу, или группу похожих молекул легко оценить, что в работе и сделано. А проследить влияние среды, многих одновременно идущих разнородных реакций - это немного антиэкспериментально.
Поэтому мне и кажется, что ответ найдут реверсивными квантовыми вычислениями :)-
Согласен, верное замечание. Но с другой стороны были же такие исследования и весьма обнадеживающие результаты показали, даже в РФ с нашим мизерным финансированием. По сути бери аппатитовую подложку и синтезируй на ней цепочки, проверяй их активность при добавлении реагентов гликогенеза, облучай УФ периодично, смотри где пошел процесс, а он же пойдет, реагенты копеешные, только анализаторы нужны. Уже был бы прорыв.
-
-
-
Я честно не понимаю почему упрямо пытаются спуститься от молекул вниз вместо того чтобы идти от минералов вверх.
@
Вот-вот, прямо в точку! Точнее, в линию. ))) Линию энергетического раздела между неживым и живым. Неживая зона началась прямо от большого взрыва, и далее понеслась эстафета превращений, идущая с расходом энергии. Частицы – атомы: 10*6 ЭВ / атомы – молекулы: 10 ЭВ / молекулы – супрамол.системы: 1 ЭВ / супрасистемы – биосистемы: 0 (ноль) / а далее бегуны понесли эстафетную палочку только на энергии извне.
На этой линии материя попала между УФ-молотом и минеральной наковальней. Конечно, только тут и могли выковаться и пре/ и а/ биотические структуры. А почему идут сверху вниз? Генетика - это биохимия, рибозимы - это машинки, в которые играют как раз генетики, им западло и просто не по уму заезжать в область классической химии и совсем уж тошно всматриваться во всякие там супрамолекулярные ансамбли...
Оно ж не просто так появился навык соображать на троих. Химик, генетик и энергетик... Правда, Высоцкий предупреждал: "Часто, разлив по сту семьдесят граммов на брата, Даже не знаешь, куда на ночлег попадёшь..."
Ну, за абиогенез!-
как раз классическая химия работает в растворе как и пытаются делать, а вот химия гетерогенных процессов идет в адсорбционном слое кристаллов, где свойства отличаются и реакции по другому идут. Им нужны химики коллоидщики, кристаллохимики в помощь, плюс конечно геохимики. А пока они будут ставить синтезы в пробирке так дело и не пойдет.
-
Согласен, классическая химия растворов тут не подходит. Лучше было бы сказать про специалистов в гетерогенно-каталитической химии. Но как обозначить их одним термином? Если "гетерогенщик" ещё можно произнести, то "катализщик" уж совсем не звучит. Длинные посты - это ужас, где эти говоруны столько времени берут?
Задача абиогенеза пока видится многим как биологическая. Пора уже увидеть её в до-биологической зоне, причем, в период жесткой солнечной радиации, когда ещё не было ни атмосферы, ни озонового слоя, и под эту обстановку подбирать узких специалистов. То есть, перекомпозировать задачу.-
да в этом тоже проблема что работы выполняют или чистые молекулярщики из всяких институтов оргхимии или напротив геохимики, и совсем нет совместных проектов
-
Чтобы были совместные многодисциплинарные работы, нужен единый руководящий центр - АН РФ. Однако, она по закону превращена в экспертное сообщество. А теперь, с крупными разрывами в геополитике, не до прорывов во второстепенных научных вопросах. Так оно и пойдёт: все прорывы будут там, а здесь, в лучшем случае, будут поддерживать тех, кто способен понять, о чём именно пишут там.
-
-
-
-
-
-
-
-
Да не нужно водой ничего размывать она все реакции вспять обращает, нужен УФ который будет разрушать постоянно днем все что нереплицируемое до новых кирпичиков. Не нужно менять ни рН ни соли, все есть на минеральной матрице и так. Замечу что скорее всего абиогенез шел на земле когда океанов как таковых вообще не было, зарождающиеся под действием приливных сил Луны конвекционные ячейки только начинали выделять воду из недр, на границах прото кратонов, везде парило, но океанов не было, максимум лужи, так что и о приливах говорить не приходится. Океаны наполнялись долго а жизнь к появлению первых кратонов уже зародилась судя по времени происхождения. Так что о приливах и океанах со всех точек зрения стоит забыть.
-


-
Не смотрел исходную публикацию, но в тексте этой статьи -
достаточно добавить в исходную смесь один-единственный активированный гексамер
Достаточно - но не обязательно именно один.-
Как я понимаю из оригинального текста статьи, фактически добавляли только один, именно нужный, гексамер: "Therefore, the inclusion of one defined RNA hexamer together with the required triplet substrates enabled selfsynthesis of the full length QT45 (+) strand by the QT45 (+) ribozyme on a QT45 (-) template (Fig. 4B)". Но, может, Александр внесёт ясность.
-
-
Проблема в том, что другие олигомеры могут мешать репликации, налипая на другие участки QT4, и мешая ему, тем самым, свернуться правильным образом. То есть, интересно было бы попробовать использовать в начальном растворе, например, смесь триплетов и гексометов. В любом случае, я согласен, что это очень интересный результат. Надеюсь, данная группа на нём не остановится, а будет постепенно продвигаться к экспериментам с большей реалистичностью начальных условий.
Если таких экспериментов не проводится, то с чем это может быть связано. Вроде бы РНК также могут объединятся нековалентно, как и белки.
-
Генетический код - это сильно позже. Отличия в нём триплетов по одному нуклеотиду - проявление избыточности для предохранения от мутаций в аминокислотной последовательности. А пока что в "первородной смеси" исключительно моно-, олиго и полинуклеотиды РНК в разных комбинациях и пропорциях. Где-то, где преобладали триплеты, "эволюция" шла быстрее.
Глобальная проблема в другом. Авторы статьи использовали D-рибонуклеотиды. И где-то на Земле или в Космосе надо найти такое место, где рибозы абиогенно образуются не в виде рацемической смеси изомеров, а исключительно в виде D-изомеров. Пока о таких местах и условиях все молчат.-
Насколько помню, РНК цепочки из нуклеотидов со смешанной хиральностью тоже вполне себе образуют вторичные укладки.
-
-
-
Кое-что проверили. https://www.nature.com/articles/nature13900
The D-RNA enzyme, consisting of 83 nucleotides, catalyzes the joining of L-mono- or oligonucleotide substrates on a complementary L-RNA template, and similarly for the L-enzyme with D-substrates and a D-template.
Но гетерохиральные цепи вряд ли могут что-то полноценно копировать, там и проверять нечего.
-
-
-
-
-
Никто не молчит на органоминеральной матрице образуются именно стерические изомеры, именно за счет реакции на поверхности а не в растворе и связанных с этим стерических затруднений.
-
И на какого же сорта органоминеральной матрице можно отгомохиралить хотя бы 2-компнентную смесь, например, фуранозы и аминокислоты?
-
на аппатитовой матрице D-рибоза получается селективно, да и прочие правовращающие сахара. Тоже и по аминокислотам были исследования. Не надо делать в колбе рацематы а потом героически разделять, надо сразу синтезировать чистые. Ну и про УФ писали что он разрушает активнее определенные изомеры. Про синтез Сазерленда писали, что даже из смесей получались оптически чистые при исходном избытке 1% одного из изомеров. Материалов масса на эту тему. Надо просто сесть и изучить литературу.
-
-
+ наверняка есть какие-то катализаторы процессов, + возможно две РНК действуя совместно могут решать эту задачу эффективней, если решают кратно эффективней, то вот и сразу заинтересованность в образовании пены, как прототипа будущей мембраны.
-
Не вулканов, а окраин формирующихся кратонов, которые постоянно выделяли пар, СО2, Н2. Луна приливами разогревала мантию и запустила около 40 конвекционных ячеек которые дали первичные кратоны в итоге, то есть вся земля была условно разбита на 40 больших ячеек в которых примерно половина периметра постоянно дегазировалась без всяких экстримов типа вулканов, которых тогда и не было в принципе.
-
я условно. На элементах была классная статья про эволюцию планеты, и что первый миллиард лет была намного более сложная система теплообмена недр с поверхностью, чем в последние 3 ярда лет.
-
Да я помню статью, да и целый учебник есть на эту тему, возможно я его там в коментах упоминал, просто употребление термина "вулкан" сразу дает неверную предпосылку что уже были континенты, океаны и где то там в брызгах лавы и сернистого газа зарождалась жизнь, это и выглядит бредово и создает имидж безумных ученых. А по сути ни континентов ни океанов ни вулканов. Соответсвенно когда почти 10% поверхности земли это мегареактор вероятность совсем не выглядит недостижимой.
-
-
См. "Полный провал гипотезы самозарождения программируемого синтеза белка, или обнадёживающая новость теории абиогенеза?"
https://dzen.ru/a/abhby31-Q1HyJuu1
-
-
можно пояснить на конкретном примере, в чём "статья-опровержение" неправа? Для меня, как дилетанта, это не совсем очевидно (а разобраться хочется) + на других площадках точно также подвергают сомнению революционность открытия. Заранее благодарю.
-
можно пояснить на конкретном примере, в чём "статья-опровержение" неправа?
Прежде всего в том, где она размещена. Правила элементарной цифровой гигиены не позволяют так вот запросто и бесплатно копаться в записях, размещаемых в мусорном ведре общественного туалета.
Если ВАм это так важно - озаботьтесь тем, чтобы извлечь её оттуда, отмыть и поместить на более приличный хостинг. Иначе - увольте.на других площадках точно также подвергают сомнению революционность открытия.
Начать следует с того, что это открытие никто и не полагал каким-то глобально-финально-революционным в проблеме абиогенеза. Оно может быть прорывным только с позиций современного уровня знаний и методов, и этим и ценно. А путь в тысячу ли, как известно, начинается с одноо шага. И ими - отдельными шагами - в общем, продолжается.
-
-
-
Готов спорить, что статью не читали, в противном случае комментарий был бы иной. Ну а если для вас комментарий рецензента биохимика из Университета Алькалы, специалиста в области химической эволюции и происхождения жизни, метаболомики, геобиологии и органической геохимии, профессора С. Сальвана - это фуфло, то видимо вы более крупный специалист.
ну так докажите, опровергните хоть один тезис критиков.-
То что это мусор понятно даже из названия, адекватный человек такого не напишет. А с вами спорить - себя не уважать.
-
Из названия сайта. На модерируемом мной форуме, на основе многолетнего опыта, просто ввёл запрет публикации авторами ссылок на мусорные сайты, включая dzen.ru
Предлагаю редакторам Элементов сделать то же самое.
Вот мой перечень для затравки:
/narod.ru/
/ok.ru/
/dzen.ru/
/utro.ru/
/aif.ru/
/overclockers.ru/
/aftershock.news/
/ura.news/-
-
-
-
Она в интернете в любом случае не будет стерильной. Речь тут только о том, что не следует жрать с помойки, ибо ДГКП, сальмонеллы, клебсиеллы, клостридии и энтеровирусы не полезны для иммунитета в любом случае.
-
Сколь часто на Элементах в комментариях дают ссылки на перечисленные вами странички? Ничтожно редко.
А narod.ru - всего навсего старый хостинг, где любой мог бесплатно сделать свой сайт на любую тему.
"не следует жрать с помойки" - читатели Элементов настолько неразборчивы и глупы? )-
Сколь часто на Элементах в комментариях дают ссылки на перечисленные вами странички? Ничтожно редко.
Ну вот выше уже один случай. Дальше - может и больше быть.А narod.ru - всего навсего старый хостинг, где любой мог бесплатно сделать свой сайт на любую тему.
Полный зловредов (вирусни и троянов)."не следует жрать с помойки" - читатели Элементов настолько неразборчивы и глупы?
Судя по комментариям перешедших по ссылке на Дзен и зачем-то разбиравших выложенный там бред - да.
-
-
-
-
-
-
Edoardo Gianni, Samantha L. Y. Kwok, Christopher J. K. Wan, Kevin Goeij, Bryce E. Clifton, Enrico S. Colizzi, James Attwater, Philipp HolligerНе вижу в авторах статьи А. Маркова. Он лишь дал обзор статьи перечисленных авторов. Вам следует высказывать свои претензии непосредственно Edoardo Gianni, контакты в свободном доступе. :)
абиогенез, самозарождение жизни, происхождение жизни, биосинтез белка, эволюция, дарвинизм, атеизм, наука, лженаука, Александр Марков
Первые пять слов — научные термины. Последние пять — нет. Это сигнал: текст написан не для того, чтобы разобрать конкретную статью Singh et al., а для того, чтобы вписать её в заранее готовый нарратив. Это не просто риторика — идеологическая рамка в начале текста создаёт конкретные методологические последствия.
2. Ad hominem вместо аргументов
Александр Марков атакуется лично: «специалист по ископаемым морским ежам», «слепая вера», «позорит науку». Это риторика, не критика. Специализация учёного не отменяет содержание его аргументов.
3. Цитата Кунина вырвана из контекста
Кунин описывает сложность проблемы, но нигде не заключает, что абиогенез невозможен или является «псевдонаукой» — он продолжает активно работать в этой области.
4. Аналогии подменяют доказательства
«Принтер под градом», «корона на свалке» — образные, но научно пустые конструкции. Они апеллируют к интуиции, а не к химической логике.
5. Вывод «абиогенез — псевдонаука» не следует из приведённых данных
Нерешённость проблемы ≠ псевдонаука. Псевдонаука — это когда теория не допускает опровержения. Абиогенез — область, где активно ставятся эксперименты, формулируются проверяемые гипотезы и публично признаются неудачи. Именно это и описывает автор, сам того не замечая.
Текст написан не как научная рецензия, а как полемический манифест. Сама по себе полемика — законный жанр. Проблема в том, что автор представляет её как объективный научный анализ, не раскрывая своей исходной позиции явно.
-
Edoardo Gianni, Samantha L. Y. Kwok, Christopher J. K. Wan, Kevin Goeij, Bryce E. Clifton, Enrico S. Colizzi, James Attwater, Philipp Holliger. A small polymerase ribozyme that can synthesize itself and its complementary strand
И это все недочёты? Ни к одному из приведённых аргументов, как я понимаю, претензий нет.-
Исправьте свои недочёты — уберите ad hominem, восстановите контекст Кунина, уберите «атеизм» и «дарвинизм» из ключевых слов, не используйте пустые аналогии. Что останется?
Останется ровно одно: наука движется к пониманию происхождения жизни — медленно, через промежуточные результаты и новые гипотезы. Это и есть научный метод: каждый эксперимент сужает пространство незнания и точнее формулирует следующий вопрос. И чем дальше она движется — тем менее нуждается в Боге для объяснения наблюдаемых явлений природы.-
-
Вы правы что Бог никуда не девается — как персонаж. Но его место в объяснении мира за последние 200 лет изменилось радикально. До Дарвина Бог объяснял всё: болезни, молнии, виды животных, происхождение человека. После Дарвина — чуть меньше. После Пастера — ещё меньше. После Хаббла Бог перестал быть нужен для объяснения устройства вселенной. После Уотсона и Крика — для объяснения наследственности. Это и есть смена символического порядка: Бог не исчезает — но перестаёт быть объяснением и уходит за кулисы отыграв свой номер.
-
Вся физика продолжает существовать в какой-то внешней среде. Вон теории струн и т.п. пытаются объяснить всё наблюдаемое, но в какой среде они сами существуют?
Если есть формула всего (допустима сама возможность её существования), то вселенной априори не существует, вся наша жизнь просто завихрения в чистой логике.
Вот тут постоянно упоминают мультивселенную, а ведь условный кардинал этой концепции не превышает кардинал концепции того, что нас со вселенной создали из другого мира. Мы сами это постоянно делаем, даже я делал несколько раз, поэтому исходя уже из практических соображений мультивселенная кажется менее вероятной концепцией.-
субатомная физика и правда не совсем «настоящая» физика, её в некотором смысле можно считать «завихрениями логики» (и всякие мультивселенные тоже)
«настоящая» физика заканчивается там, где заканчивается эксперимент (а эксперименты на адронном коллайдере содержат между эффектами «слишком много» математики :3)
это и правда кризисное состояние физики (и даже науки в целом), мы по сути наделяем статусом физического существования математические конструкции (т.е., какая-нибудь струна или волновая функция электрона - это не какое-то там приближение или аппроксимация физического объекта, а буквально сам физический объект, который мы и исследуем)
проблема отравления естественных наук платонизмом существует (а с успехом машинного обучения усугубляется многократно), она известна, обсуждаема, но что с этим делать - пока неясно
пока просто улыбаемся и машем :3
https://chat.deepseek.com/share/brpgz5pqxmp66ej34l
https://chat.qwen.ai/s/t_781e5b0d-774c-4b85-ba0c-6aca681ba832 -
субатомная физика и правда не совсем «настоящая» физика
это не какое-то там приближение или аппроксимация физического объекта
Ну это какое-то совсем дикое понимание физики, на уровне 19ого века.
Наоборот, волновые функции первичны.-
так об этой проблеме и речь в моём спиче, волновая функция (математический объект) становится физическим объектом
это методологический провал и как раз средневековый платонизм под соусом новомодного «физического нигилизма» (неоплатонизма) :3
язык науки (математика) стал первичнее (в онтологическом смысле) предмета науки (объективной реальности), карта и территория поменялись местами :3
ещё чуть-чуть, и мир будет существовать только потому, что я его наблюдаю (я, конкретный я, конкретный солипсист, поддерживающий всю вселенную вокруг своим осознаванием её существования) :3
(на самом деле и такие концепции уже в разработке в науке на самых серьёзных щах)
…в социально-экономическом плане это всё следствие капитализма, маркетинга, форса радикального индивидуализма, именно он гнёт мозги в сторону «магического мышления второго порядка» - «я сам ответственнен за свой успех, системные эффекты вторичны, объективная реальность менее важна, чем субъективная» :3
https://chat.qwen.ai/s/t_28cc841c-2cb2-4c23-a77f-2ed73a014eaf -
это методологический провал и как раз средневековый платонизм под соусом новомодного «физического нигилизма» (неоплатонизма) :3
Нет конечно. Нужно понимать, что она первична, именно она является средой где существует понятная нам физика. А понятна нам эта физика, потому, что наш мозг заточен для того, чтобы бегать по Африке и собирать дохлых жирафов.
Если бы скажем вы бы были дельфином, возможно вам бы вам волновая функция казалась намного более естественной и понятной.
Представьте себе мудреца живущего на вершине горы и наблюдающие внизу синюю равнину с белыми полосками. Всю жизнь наблюдал, учился сопоставлять белые полоски с погодой, и воспринимал мир, как гору и синюю равнину. А потом спустился вниз и увидел, что это море с волнами и вообще непонятным поведением, которого никогда не видел. Он может ваши аргументы повторить.
Нужно уметь смотреть на мир с разных точек зрения, выбирая наиболее подходящую под контекст, откидывать текущие шаблоны и набирать новые, соответствующие ситуации.-
нужно понимать методологическую проблему, а не спорить за церковь математики (платонизма, магического мышления) :3
-
Методологическая проблема это обезьяньи мозги.
Многие любят почему-то абсолютизировать интеллект, типа есть ли интеллект у кого-то ещё. А это просто инструмент возникший для работы в определённых условиях, он неуниверсален.
Типа абстрагирование позволяет несколько избавиться от яблок и груш, но в чистом виде это требует нехилых усилий и приличной головной боли.-
да, обезьяньи мозги склонны к магическому мышлению, это первичный субстрат нашего мышления, и наука (методология объективизации знаний) - это уже довольно нетривиальная надстройка над ним, но некоторые слабые духом скатываются обратно к платонической магии и солипсизму :3
(ещё и бравируют этим, строят теории превосходства на базе этой своей "особенности" :3)
-
-
-
-
-
-
-
-
-
-
-
Аналогии подменяют доказательства
«Принтер под градом», «корона на свалке» — образные, но научно пустые конструкции. Они апеллируют к интуиции, а не к химической логике.
Не совсем согласен. Например "парадокс миллиона обезьян" вполне применим в научном диспуте и активно используется. Такие примеры хорошо отрезвляют многие фантазии.
А вот обзор обсуждаемой публикации, хоть Марков и присутсвует, но уже не как главный герой: "Мы очень близки к полному краху гипотезы “мира РНК” – крупнейшей аферы в науке XXI века" https://dzen.ru/a/acMRciSeQkWAF2FC
-
Нет тут никакого особого открытия. Эта работа интересная, показывает новую методологию, но реально ни на какие ранее неизвестные вопросы не отвечает.
QT45 не является искомым репликатором. Скорее всего должно удачно сложиться несколько молекул РНК, причём параллельно должен идти ещё другой процесс завязанный на солнечное изучение.-
Если есть какой-то работающий репликатор, то сделать из него "искомый" — дело техники. А солнечное излучение — процесс не случайный, поэтому в формулах расчёта вероятностей это всегда константа. Кроме того, всегда есть места, где это излучение максимально, равно 0 и принимает промежуточные значения.
-
1. Нет, не дело техники. Этот формат репликаторов очень маловероятно, что имеет отношение к вопросу появления жизни. Kordum несёт много эмоций, но по существу постом ниже правильно сказал.
Нужно решать проблему хиральности - вот это было бы открытием.
Нужно многократно повышать эффективность.
Скорее всего нужно смотреть репликацию в целой группе относительно простых процессов протекающих одновременно. Не репликатор как таковой, а сеть реакций.
2. Скорее всего процесс был завязан на солнечное излучение в присутствии сульфата цинка, это часть уравнения, увязав которую с репликаторами можно было бы говорить о чём-то особо интересном.
Как мне кажется, решение будет найдено когда дооптимизируют квантовые вычисления и просто реверсивным образом подберут соответствующие реакции.-
Kordum несёт много эмоций, но по существу постом ниже правильно сказал.
Спасибо за честность. Вы только что подтвердили оба моих тезиса: текст креациониста эмоциональный — значит не научный. А «прав по существу» в том единственном пункте, с которым никто никогда не спорил — эксперименты неполны. Это не защита статьи. Это её некролог.-
текст креациониста эмоциональный — значит не научный
Эмоциональность текста никак не может определять его научность или не научность. Важны только факты, а способ подачи - дело вкуса.-
Может. Наличие эмоций делает текст бессмысленным с точки зрения подачи фактов и возможности их осмысления.
Эмоциональное мышление намного сильнее аналитического, в него легче упасть, а при злоупотреблении это происходит необратимо. При этом оно примитивно.
Поэтому первое правило когнитивной гигиены - любой текст с эмоциями считается мусором, как пропаганда, где автор пытается тобой гнусно манипулировать.-
Вы сейчас написали про удобство чтения. Тут согласен - хочешь чтобы тебя поняли или даже просто прочитали, пиши определенным образом. Но! Даже написанный эмоционально, с оскорблениями и максимально токсично текст - не перестаёт быть научным, если научна его система аргументации. Иными словами, форма не отменяет содержания.
К тому же, есть подозрение, что добрая часть науки XIX века была тем еще токсичным болотом, где переход на личности был частью культуры))-
Это не вопрос удобства чтения. Как только у нас в голове включают эмоции - наша рассудочная способность стремительно падает. Мы теряем способность к анализу, физиологически, в прямом смысле этого слова.
Поэтому любые эмоции в материале, который мы потребляем не ради эмоций - показатель мусорности, лживости материала.
Вкрапления фактов нужно воспринимать, как способ втереться в доверие, чтобы вам зашла основная ложь. Это касается науки, политики, экономики, всего.
В 19ом веке и начале 20ого наука была совсем не такая как сейчас, учёные были интеллектуальной элитой своего времени. Именно в научных трудах эмоций мало было, перепалки были вне их. Эмоции в трудах этнографов победили, поэтому вся западная Европа в этом плане упала в лженауку, и не смогла преодолеть свою дикость в этом плане.
-
-
-
-
-
-
-
-
Даже если и на десятки млн лет, это ничего принципиально не меняет. Хотя я слышал оценку 100 - 300 млн лет, которые были у абиогенеза с момента, когда Земля стала пригодной для него, и до момента, когда появляются намёки на существование жизни.
-
-
Я имел в виду, что 4.4 млрд лет назад поверхность Земли уже точно не была расплавленной, судя по сохранившимся с того времени породам, а 4.1 млрд лет назад вроде бы есть намёки на существование жизни, и это 300 млн лет. Точно же установлено существование жизни для времени 3.9 млрд лет назад, и это ещё 200 млн лет.
-
Точно же установлено существование жизни для времени 3.9 млрд лет назад
Сомнительное заявление. У нас с тех пор только цирконы остались, из них не слишком много чего можно вытащить.-
правильно вся поверхность тогда была постепенно переработана, соответсвенно жизнь вынуждена была постоянно перебираться на новые места, и следов первой жизни найти невозможно в принципе, по сути пока первые кратоны не превратились в протоконтиненты и океан не заполнил все пространство между ними, никаких шансов на сохранение следов жизни не было.
-
Согласен. Слышал когда-то такие временные оценки, но сейчас посмотрел и увидел, что все эти временные интервалы сильно размыты. Однако это даже расширяет время возможного абиогенеза чуть ли ни до миллиарда лет, что только увеличивает вероятность зарождения жизни таким способом. Хотя, на мой взгляд, в свете этой статьи, и нескольких миллионов лет достаточно, даже с учётом всех проблем, связанных с их экспериментом.
-
-
-
-
-
Археи такие пошли мы нафиг :) Ведь они скорее всего длительное время доминировали. А ведь на заре, вполне вероятно, что были и другие варианты прокариот.
Но надо понимать, что от возникновения планеты до первых следов у нас около миллиарда лет. Сколько то сотен миллионов ушло на релаксацию планеты, но всё-равно, первые +-600М когда жизнь формировалась нам не видны.-
-
Источник? Заявление, что археи появились из бактерий выглядит довольно странно. Нигде не видел таких статей.
-
по моему я понял об чем речь, была такая статья но там если память не изменяет говорилось о сеточной эволюции когда археи заимствовали что то бактерий и это автоматом их омолаживало. Как мне показалось из той статьи авторы скорее делали вывод что ранняя эволюция архей и прокариот была слишком сложной чтобы достоверно установить кто раньше.
-
-
Вот, например, одна из самых свежих статей, в которой говорится, что ЛУКА был ацетагеном, а отнюдь не метаногеном и обладал клеточной стенкой из пептидогликана: https://www.nature.com/articles/s41559-024-02461-1
Понятно, что чёткого разделения провести невозможно, так как разделение бактерий и архей это, по определению, всегда корневая точка ветвления после LUCA.-
Мы эту статью обсуждали.
1. Бывают археи ацетогены. За столько лет структура их сообществ могла сильно измениться, поэтому это аргументом не является.
2. Лука не был конкретным организмом с точки зрения генома, это была популяция, в которой существовал дрейф генов. Т.е.возможно, что физическая капля которая делилась и дала все нынеживущие организмы таки была, но генный материал, который дошёл до наших дней в заметном количестве был не в ней на тот момент.
3. Они пишут про пути пептидогликана, но про саму мембрану точно не пишут.
ИИшка выдала такое:
У бактерий пептидогликан — обычная, хорошо аннотированная и широко распространенная система. У архей клеточные оболочки гораздо более разнородны: S-layer, псевдомуреин у части метаногенов, разные липидные архитектуры. При этом авторы сами признают ограничение KEGG-подхода: некоторые древние и широко распространенные семейства могут быть разбиты на несколько KO, которые выглядят как таксономически ограниченные и потому “кажутся” более поздними. Для этого они добавили более грубый анализ COG, но это не полностью снимает проблему. То есть методологически бактериоподобные, лучше аннотированные пути действительно могут восстанавливаться легче, чем архейные инновации или древние архейные варианты оболочки.-
Археи - ацетагены появились сравнительно поздно, скорее всего это ГПГ. Пептидогликан это не про мембрану, а про клеточную стенку. На самом деле, это во многом вопрос определения, если что-то выглядит как кошка, и мяукает как кошка, то, на мой взгляд, как завещал нам старик Оккам, это логично и считать кошкой, пока не доказано обратное. Но я не настаиваю, в принципе, вопрос гораздо менее принципиальный, чем странные нескладухи с молекулярными часами.)
-
Вообще, если пофантазировать по некоторым другим примерам эволюции, Лука, как общий предок архей и бактерий, мог бы иметь какую-нибудь странную конструкцию вместо текущих мембран и стенок, в связи с тем, что на ранней стадии непонятно что было делать с транспортом веществ, до развития соответствующих белков.
Типа может они, на уровне клеток изначально прикреплялись к субстрату и от него получали питательные вещества контактируя цитоплазмой напрямую, а от остального пространства защищались всем подряд.
Можно пофантазировать, что иногда они отрывались от субстрата, в режиме голода перелетали на новое место и там прикреплялись заново, и вели такой осёдлый образ жизни, до тех пор, пока не наработали мембранные белки и не упростили конструкцию оболочки до бактериальной и архейной формы.-
Ну логично предположить что "мембрана" изначально была просто длинными молекулами прикрепленными к субстрату на котором она и синтезировалась, и вообще была побочным продуктом, соответственно функция защиты от размывания у нее была число случайная, защитить нижний синтезирующий слой протоРНК при неизбежных брызгах или осадках. Конечно она была из чего попало более менее длинного. Логично что перенос у нее был пассивный, попала вода или органика пошла диффузия между молекул. И только когда пошел отбор на защитные свойства она стала улучшаться. Понятно и очевидно что постоянная работа кристалла в виде связанной АМФ-АДФ-АТФ приведет в итоге к ослаблению и разрушению связи и откалыванию участков полифосфатов от основного кристалла. Соответсвенно периодически микрокристаллы со всей обсадкой откалывались и переносились влагой или ветром, и такой кристалл покрытый синтезирующей органикой условно мог стать прообразом клетки. Насколько помню до сих пор ряд видов одноклеточных запасает энергию в виде полифосфатов внутри клетки. Чем не реликт прошлого?
-
-
-
-
-
-
-
Причём наверняка существовало обилие разных типов прокариот, наверняка были какие-то здоровые и очень необычные. Но не выдержавшие наступление воды, как позже уже другие формы не выдерживали наступление кислорода.
А предки архей и бактерий были какими-нибудь неудачниками, с краю, которых спихнули в воду, но те выжили :)-
Тут самое интересное что правильная теория абиогенеза просто обязывает появится вирусам, они неотъемлимая часть абиогенеза, изначально возникшая как конкурирующие не целевые реакции, не влившиеся в протоклетку. Отсюда сразу снимается известная проблема откуда они появились сразу и такие сложные. Ну а то что прокариоты были разные по определению понятно, не могли быть они одинаковые. Теория что это неудачники на самом деле имеет место, наверняка самые удачливые заняли козырные мста где не надо было прятаться под шубу из липидов те же например поры минералов, но оказавшись в наступавшей воде они все разумеется погибли, а неудачники на которых лило и брызгало и смывало в итоге и выжили. Вполне правдоподобная гипотеза.
-
https://www.nature.com/articles/s41598-020-58060-0 "There is a reasonable hope to find a functioning replicase ribozyme longer than 40–60 nt.", которые всё равно меньше необходимого для многократного зарождения жизни в масштабах галактики, если это - чисто случайный _пуассонов_процесс:
"Minimum RNA length must be lmin = 21, 27 and 32 to expect one abiogenesis event for a survey of a single star (lgN* = 0), a galaxy (lgN* = 11), and the observable universe (lgN* = 22), respectively."
-
Насколько я понял, он считает, что вероятность возникновения нужной цепочки до 20 нуклеотидов на отдельно взятой планете приближается к 1 ("On the other hand, lmin must be shorter than ~20 nucleotides for the abiogenesis probability close to unity on a terrestrial planet"), при этом вероятности и времена рассчитывает исходя из того, что за раз к цепочке присоединяется 1 нуклеотид (формула 1: x'(l+1)=k*x(l)-k*x(l+1)). Здесь же присоединяются триплеты, и если считать в них, то длина всей цепочки всего 15 триплетов, что на 5 меньше минимально необходимой длины для вероятности ~ 1. Кроме того, в другом месте статьи он произвольно полагает, что из всех возможных последовательностей длиной в 40 нт только 10^4 (10000) годны для репликации ("Considering an example with lmin = 40, there are 4^40 ~ 10^24 possible sequences of 40-mers, and perhaps Nac = 10^4 sequences out of them may have a replicase activity"). Здесь же показано, что из всех последовательностей длиной 45 нт к репликации годны ~10^11, что увеличивает вероятность случайной самосборки ещё на несколько порядков.
-
Насколько я понял, он считает, что вероятность возникновения нужной цепочки до 20 нуклеотидов на отдельно взятой планете приближается к 1
Так их всего 1 099 511 627 776 вариантов такой длины! Их даже не на планете, а в одной цистерне/болоте реагентов может возникнуть не одна!
Даже всех вариантов олигонуклеотидов длиной до 30 звеньев будет меньше 1 моля. Но вот примерно с этого момента в реальности всё уже становится сильно сложнее - как в той задаче про зарастание пруда кувшинками.-
Я так и понял, но, мне кажется, это не отменяет сказанного мною выше про триплеты, увеличивающие скорость роста цепочек, и про ~10^11 способных к репликации цепочек длиной в 45 нт, увеличивающих вероятность удачного исхода за заданное время.
-
Причём тут скорость роста и длина конкретных цепочек? В оценке Тотани - только минимальная длина всей КОНСЕРВАТИВНОЙ последовательности. Она даже может где-то прерываться какими-то спейсерами какой-то длины, если это нужно или наоборот - не важно. главное - что она нужна СРАЗУ ВСЯ ТАКАЯ для запуска цикла Дарвина-Эйгена без вырождения.
-
В оценке Тотани - только минимальная длина всей КОНСЕРВАТИВНОЙ последовательности.
Ну да, и он оценил её в 40 — 50 нт, что полностью подтверждено данным опытом. А дальше он начинает оценивать вероятность возникновения последовательности такой длины, и у него получается, что практически со 100% вероятностью на Земле могут появиться только цепочки до 20 нт, а рабочие цепочки 40 — 50 нт появляются с вероятностью сильно меньшей. Но при оценке этих вероятностей, если я его правильно понял (могу и ошибаться), он исходит из того, что:
1.) Нуклеотиды добавляются по одному, а время ограничено, т. к. внешние факторы постепенно разрушают цепочку. — Но если они добавляются по 3, как в данном опыте, то и время роста цепочки сокращается в 3 раза.
2.) Для цепочки длиной 40 он предполагает 10^4 удачных последовательностей, что соответствует 10^7 удачных последовательностей для цепочки длиной 45. В данном же опыте удачных последовательностей ~ 10^11, что на 4 порядка или в 10000 раз повышает вероятность удачного исхода по сравнению с его прогнозом.-
у него получается, что практически со 100% вероятностью на Земле могут появиться только цепочки до 20 нт
На мой взгляд, вполне могут и больше - до 24 (верхний размер стандартного ДНК-праймера для ПЦР) уж точно. Да и среди существующих геномов сколь-нибудь уникально-специфичные последовательности тоже начинаются только примерно с этой длины. Надёжно - от 30. И вот где-то в этом диапазоне ~25~35 нуклеотидов и лежат пороги "невероятности" самосборки последовательности в масштабах звёздно-планетных систем. -
где-то в этом диапазоне ~25~35 нуклеотидов и лежат пороги "невероятности" самосборки последовательности в масштабах звёздно-планетных систем.
А откуда берутся эти пороги невероятности? Из простой комбинаторики без дополнительных ограничений эти пороги не следуют. В статье указывается на временное ограничение и делается вывод, что при прирастании 1 нуклеотида за раз успеет склеиться до 20 нуклеотидов. Здесь они приклеиваются по 3, а значит за то же время РНК может вырасти до 60, чего более чем достаточно. Значит, должны быть ещё какие-то ограничения на рост.
-
-
-
-
-
------------------------------------------------
Это доказывает, что господь-бог мог создать жизнь на Земле занимаясь случайным перебором полимераз.
-
Репликатор, который по сути представляет собой молекулярный мусор, т.к. для живой клетки непригоден и в её формировании не участвует. Настоящие рибозимы реплицироваться не способны из-за шпилек, наконец-то это признали, т.е. мир рибозимов это утопия. К тому же репликация идёт только под руководством высококвалифицированных химиков. Так что это статья - очередная профанация и не более того.
-
Репликатор, который по сути представляет собой молекулярный мусор, т.к. для живой клетки непригоден и в её формировании не участвует.
Не мусор, ибо может в принципе очень даже участвовать - как некая первичная РНК-полимераза. Равно как и рибозимная активность вообще - например в процессах (само)сплайсинга...
-
1. Реплицирует себя лишь в крайне специфических условиях (эвтектический лёд, температура около −7 °C, щелочная среда, обязательное присутствие ионов Mg²⁺ в строго определённой концентрации) и при отсутствии прочих компонентов, предполагаемых в моделях «первичного бульона».
2. В качестве субстратов использует не отдельные нуклеотиды, а тринуклеотидтрифосфаты (активированные триплеты), причём в строго определённых концентрациях.
3. Процесс протекает в два этапа, требующих различных условий: для синтеза одной цепи необходима смесь из 64 триплетов, а для второй – 13 строго комплементарных триплетов и один гексамер (все компоненты должны находиться в активированном состоянии).
4. Обладает невысокой точностью репликации: на первом этапе получено 10,9 % полностью идентичных молекул, на втором – 43,4 %. Итоговая точность составляет 0,109 × 0,434 = 0,0473 (4,73 %), то есть из 1000 синтезированных цепей корректными оказываются около 47. При этом функциональность рибозима чувствительна к мутациям: проведённые авторами тесты (однонуклеотидные замены, делеции и двойные замены) показали, что подавляющее большинство изменений существенно снижает активность, особенно в каталитических участках структуры.
5. Характеризуется крайне низким выходом продукта: около 0,24 % за 72 суток на первом этапе и 0,17 % – на втором. Итоговый выход составляет примерно 0,00041 %: около одной успешной полной репликации на 245 000 попыток за 72 дня.
6. Не является самоподдерживающейся системой: требует точного подбора условий, реагентов и контроля со стороны исследователей.
7. Не демонстрирует полноценного самовоспроизведения: для этого необходим замкнутый цикл (оригинал → комплементарная копия → новая копия и т.д.). Полученные копии не инициируют следующий цикл репликации. Более того, даже при гипотетическом запуске такого цикла накопление ошибок привело бы к быстрой потере (уже через 2–3 цикла) каталитической активности.
И это всё, на что способен самый «эффективный» самокопирующийся рибозим, отобранный из 12 триллионов случайных последовательностей. Получается, что мелкие рибозимы очень и очень слабы как по скорости, так и по точности, и при этом крайне требовательны к условиям и реагентам, а крупные, хоть и сильнее и менее «капризны», но при этом не способны к самокопированию.
Вывод вполне очевиден: «мир РНК» организмов, некогда предложенный для решения проблемы самозарождения жизни, оказался чистейшей фикцией.
Из нового обзора: Мы очень близки к полному краху гипотезы “мира РНК” – крупнейшей аферы в науке XXI века https://dzen.ru/a/acMRciSeQkWAF2FC
-
-
-
То, что ты называешь опровержением, это просто твое желание занять позицию лакановского "истерика" (не истерика в бытовом смысле, конечно) - критиковать ради того что бы выглядеть умным, а не чтобы найти истину. Критика с 4-й лакановской позиции предполагает смену символического порядка, то-есть как минимум выдвижение альтернативной гипотезы - как альтернативной претензии на истину от чего ты отказался.
-
-
-
-
Я понял, что Kordum пишет вообще и какова его позиция
https://dzen.ru/a/abhby31-Q1HyJuu1
Поэтому попросил его не маскироваться за наукообразными комментариями, тем более (температура -7) это условие из статьи Singh (тиоэфиры), а не из статьи по QT45, а «Под руководством высококвалифицированных химиков» — это верно как описание эксперимента, но логически некорректно как аргумент.
Остальные его аргументы правильно показывают, что QT45 не является демонстрацией реального абиогенеза. С этим никто не спорит, включая самих авторов.
Но из этого не следует вывод «живой системы с РНК-репликаторами не может существовать».-
То что было изначально постулировано как РНК мир скорее всего никогда и не существовало, и в этом кордум прав, не важно случайно или специально и что он думал этим доказать. О несостоятельности теории РНК мира я говорил еще в 2007 году, и именно потому что я считаю абиогенез реальным, а вот РНК-мир оторванная теория от реальных условий. Конечно она что то объясняет и даже в какой то момент была прорывной, но увы пока она не будет рассматривать всю реалистичную пребиотическую обвязку она так и останется "сферическим конем в вакууме".
-
я не жду ничего прорывного в теории абиогенеза в ближайшие 10 лет
Claude мне составил список за чем нужно следить:
Карелл (Мюнхен) - Механизм синтеза пептидов на РНК
Пауэр (Лондон) - Связь метаболизма с генетическим кодом
FEBS - Градуальная эволюция репликации
Французская группа - Совместимость всех трёх компонентов
слежу!
-
-
-
Вы отвергаете мармеладную гипотезу темной материи как ненаучную. Какова ваша альтернативная НАУЧНАЯ гипотеза? То, что ты называешь опровержением, это просто твое желание занять позицию лакановского "истерика" (не истерика в бытовом смысле, конечно) - критиковать ради того что бы выглядеть умным, а не чтобы найти истину. Критика с 4-й лакановской позиции предполагает смену символического порядка, то-есть как минимум выдвижение альтернативной гипотезы - как альтернативной претензии на истину от чего ты отказался.Согласитесь, выглядеть я буду крайне нелепо! Несогласие с моей мармеладной теорией никак не подразумевает необходимость альтернативы. Достаточно того, что мои аргументы несостоятельны.
-
Неверная аналогия. Кордум (что по киргизски переводится я увидел), умело маскирует свою позицию креациониста - он привел опровержение в конце концов: абиогенез нарушает основной закон биологии, который преподаётся в школе: "Жизнь происходит только от жизни".
NikeByNike утверздает, что: для опровержения не нужно предлагать альтернатив :) Нет связи между альтернативами и опровержением.Вы отвергаете мармеладную гипотезу темной материи как ненаучную. Какова ваша альтернативная НАУЧНАЯ гипотеза?
ГИПОТЕЗА: Темная материя - небарионная материя, а мармелад состоит из барионов!
Можно конечно схитрить и выдвинуть гипотезу: Темная материя - немармеладная материя, но это будет игра слов.Согласитесь, выглядеть я буду крайне нелепо!
Крайне нелепо, а скорее психологически некомфортно не привести очевидный контрпример, поэтому я и обращаюсь к Лакану, чтобы понять этот дискомфорт!-
ГИПОТЕЗА: Темная материя - небарионная материя, а мармелад состоит из барионов! Можно конечно схитрить и выдвинуть гипотезу: Темная материя - немармеладная материя, но это будет игра слов.
Ваша альтернатива лишь показывает, чем тёмная материя не является. Никакой самостоятельной гипотезы, никаких проверяемый фактов... Забавно, но это ровно тоже самое, что делает Кордум: он не отвечает, как возникла жизнь, но он показывает (не могу судить правильно ли), как она не могла возникнуть. Между вами, буквально, нет разницы.
-
-
Колбасит не противников теории самозарождения жизни как таковых, а противников псевдонаучных теорий, пытающихся замаскироваться под науку. Почему-то людям хочется вернуть античную идею самозарождения жизни в современную реальность. Это выглядит смешно, если учесть, что любая клетка - это высокотехнологичный запрограммированный наноробот в 1000-и раз сложнее космической ракеты.
-
Хорошо. Допустим, завтра учёные создадут в лаборатории самовоспроизводящуюся молекулярную систему из простых химических компонентов. Это изменит вашу позицию?
-
Встречный вопрос: допустим учёные создадут вечный двигатель, это изменит вашу позицию по отношению к общепринятому закону физики?
-
Вечный двигатель нарушает второй закон термодинамики — он сформулирован, математически выражен и многократно проверен. Если это аналогия с абиогенезом, тогда скажите: какой именно закон природы нарушает абиогенез — и где он сформулирован?
-
Он нарушает основной закон биологии, который преподаётся в школе: "Жизнь происходит только от жизни"
-
СОВРЕМЕННАЯ жизнь (то есть совокупность живых организмов) - экологически - как минимум последнием пару миллиадов лет - со времён кислородного события. Про происхождение самой жизни этот закон ничего не говорит и говорить не может в принципе, поскольку обращается в логический порочный круг.
-
1. А какой закон говорит о происхождении жизни? Назовите хоть один закон, который направляет физико-химические процессы в мёртвой молекулярной среде к формированию жизни.
2. Вы говорите "СОВРЕМЕННАЯ жизнь", а чем она отличается от несовременной жизни? Что, науке известна какая-то другая форма жизни? Нам известна только одна жизнь - она и современная и доисторическая, состоящая из клеток, ДНК, белков, процессов репликации, биосинтеза белка, репарации, метаболизма и пр.-
А какой закон говорит о происхождении жизни?
Никакой. И никакой не запрещает абиогенез.Назовите хоть один закон, который направляет физико-химические процессы в мёртвой молекулярной среде к формированию жизни.
Не закон. Например принцип комплиментарности. Без него вряд ли вообще что-то может получиться живое в этой вселенной.Вы говорите "СОВРЕМЕННАЯ жизнь", а чем она отличается от несовременной жизни?
Как минимум широчайшим распространением систем защиты от кислородных радикалов. До кислородного события, завершившего архейский эон, у клеток в открытой среде не было в них особой нужды.Что, науке известна какая-то другая форма жизни?
Из клеточных - две простых одного корня (бактерии и археи) и одна сложная из них получившаяся (эукариоты). Ещё неклеточные есть (вирусы, плазмиды, вироиды).-
Как минимум широчайшим распространением систем защиты от кислородных радикалов.
Вы говорить с позиции сторонника бескислородной первичной атмосферы, но это не единственная версия. В научной литературе постоянно я натыкаюсь на аргументы в пользу изначально кислородной атмосферы. Бескислородная - это требование теории абиогенеза, не более того.-
Это невозможно состав газопылевых облаков восстановительный и в таких условиях весь кислород при формировании планеты будет связан в оксиды, так что исходная атмосфера всегда восстановительная, но поскольку водород выдувается солнечным ветром в итоге атмосфера постепенно становится нейтральной. Ни на одной планете солнечной системы нет окислительной атмосферы кроме земли.
-
-
В научной литературе вы на такое не наткнетесь. Вы дзен читаете, батенька. Там кроме мусора ничего нет. Собственно, поэтому то, что вы оттуда прете, никто не хочет читать. Почитайте что-нибудь помимо кладбища нейрослопа
-
Батенька, а вы что эксперт по всей базе научной литературы, что знаете абсолютно все статьи на тему абиогенеза? Что там есть и чего нет, или просто решили сделать вброс, авось прокатит. Не прокатило. На дзене есть Никитин М. , Дробышевский С. и почти все популяризаторы атеизма, хорошо что вы сказали, что это мусор.
Профессор РАН Лев Мухин, многие годы занимавшийся проблемой первичной атмосферы Земли и газового состава метеоритов, писал: «О составе атмосферы можно судить по газам, содержащимся в земных породах и метеоритах на период времени их формирования. Интересно, что эта атмосфера неравновесна: в ней есть полностью окисленные компоненты, такие как СО2, и есть компоненты восстановленные: нет аммиака, а есть только свободный азот. Тем не менее, эти данные приводят к выводу о том, что 3,8–3,9 млрд лет назад не было никакой восстановительной атмосферы. Наблюдаются следы кислорода, который должен был образовываться во время высокотемпературных процессов и очень быстро реагировать с другими веществами. Это важно потому, что в присутствии кислорода не проходят никакие абиогенные синтезы органических соединений» [Мухин Л. М. Условия на поверхности Земли 4–4,6 млрд лет назад. Первичные синтезы // Проблемы происхождения жизни. РАН. Сборник научных статей. М.: ПИН РАН. 2009. С. 120–128].
Также и Френсис Крик писал, что ранее не был принят во внимание и тот факт, что под действием УФ-лучей происходит фотолиз паров воды (как сейчас над озоновым слоем атмосферы Земли), следовательно уровень кислорода за относительно короткий период времени поднялся бы до современного [Крик, 2002].
-
-
-
-
-
-
-
Брателло, а ты в какой школе учился, 2 класса и 3 коридора закончил? В современной школьной программе 10–11 классов (ФГОС, базовый и углублённый уровень) по теме «Возникновение жизни на Земле» креационизм изучают как донаучное представления.
В какой школе ты сумел найти такой «альтернативно углублённый» подход к биологии? В спецклассе для будущих нобелевских лауреатов по альтернативной биологии?-
Я сам учитель биологии и прекрасно знаю, что в современных школах учебниках в теме "Возникновение жизни на Земле" представлено несколько теорий, обычно их 4. А дальше сказано, что ни одна из этих теорий до сих пор не получила подтверждения. А потому каждый имеет право придерживаться любой теории, и никто не может заставить принимать ту или иную версию.
-
Я сам учитель биологии и прекрасно знаю, что в современных школах учебниках в теме "Возникновение жизни на Земле" представлено несколько теорий, обычно их 4. А дальше сказано, что ни одна из этих теорий до сих пор не получила подтверждения.
Вы сами только что сказали: школьные учебники представляют несколько теорий, и каждый вправе выбирать. Значит если завтра учёные создадут самовоспроизводящуюся молекулярную систему из простых химических компонентов — учебники изменятся, и абиогенез получит подтверждение. Вы это примете? Или окажется что ваш выбор теории не зависит от доказательств — ни от школьных учебников, ни от лабораторных результатов?
По запрету вечного двигателя в учебниках нет 4-х вариантов, поэтому ваш встречный вопрос уже не поможет уйти ответа!-
завтра учёные создадут самовоспроизводящуюся молекулярную систему
Как раз-таки выбор есть там, где нет доказательств. Поэтому авторы учебников и подчёркивают, что ни одна теория не доказана, а иначе зачем приводить все версии. А насчёт "завтра", этого точно не будет, в комментарии уже сказали, что для этого нужно много времени, по сути неограниченное количество, пока не создадут, миллионы, миллиарды,триллионы лет, не важно, в этом вопросе главное - вера.
-
-
-
-
-
-
Ого! Да вы верите в витализм - идею, распространенную среди учёных до середины 19 века. В 1828 году Фридрих Вёлер искусственно синтезировал мочевину. Этот эксперимент показал, что органические вещества можно синтезировать в лаборатории и никакой мистической силы для объяснения существования живых организмов не требуется. И что химические законы одинаковы для живой и неживой природы.
-
-
Особенно веселит, что сами моськи не имеют ровно никаких экспериментальных подтверждений собственным воззрениям. Ноль - вот что за их утверждениями "о невозможности". Ровно ноль
-
Зачем креацианистам нужны экспериментальные подтверждения собственным воззрениям?
Они говорят: «У вас нет ни одного эксперимента, где из неживого получилась бы живая клетка → значит, ваша гипотеза абиогенеза такая же ЭКСПЕРИМЕНТАЛЬНО недоказанная вера, как и наша вера в Творца».
Это позволяет перевести дискуссию из плоскости «у науки есть огромный массив косвенных данных + прогресс в моделях» в плоскость «никто ничего не доказал окончательно → выбирай веру по вкусу».
Требовать «сделайте мне живую клетку в пробирке за 2–3 года» — это как требовать от геолога «сделайте мне новые Гималаи за выходные», чтобы потом сказать «геология — лженаука».
Креационисты прекрасно знают, что такой эксперимент сейчас невозможен технически — и именно поэтому настаивают именно на нём.
Ну вы то уж должны понять!-
...но есть и другая сторона ситуации: их УЖЕ затащили на ту сторону поля, где они даже не тявкают о возникновении всей клетки сразу (слишком многие в курсе, слишком многие ржут с таких чайников, которые подобное излагают), а вынуждены именно что противостоять самой идее самореплицирующихся рибозимов в той или иной форме. А время не стоит на месте. И буквально через несколько лет число таких известных рибозимов будет увеличиваться, как сейчас - число вновь открываемых экзопланет. И знаете, каким станет их следующее, фактически последнее утверждение? Оно будет о невозможности перескока между вершинами в ландшафте приспособленности успешно самореплицирующихся рибозимов)) Разумеется, крайне ненадолго.
-
С позиции Лакана вторжение Реального сильнее всего подрывает ритуальный порядок любой религии, особенно фундаментальной. Именно поэтому каждый новый рибозим — это не просто «научный факт», для человека, привыкшего к Символическому порядку происхождения (Бог создал жизнь, всё устроено и понятно), это ощущается как jouissance — эмоциональная реакция на столкновение Символического (религиозного, упорядоченного смысла) с экспериментально наблюдаемым Реальным (эволюцией рибозимов) — интеллектуально-экзистенциальная паника, почти телесно ощущаемая каждой мышцей его организма, в тот самый момент, когда он не поддается травматическому насилию науки.
-
Ага, только все люди сектанты, и чёт мне начинает казаться, что сектанты примерно в равной степени.
И научный сектантизм (я не про вас, вы тупо сатанист), точно так же является проблемой для интеллектуальных людей, как и религиозный.-
Ты говоришь, что вера в плоскую Землю и квантовая механика — две равноценные секты. На деле они устроены совершенно по-разному.
Одна "секта" говорит: «Вот готовый, окончательный ответ. Всё уже объяснено, всё имеет смысл, случайности и непонятного нет. Просто прими это — и тебе станет спокойно».
Другая "секта" говорит: «Мы никогда не получим полного ответа. Каждый новый шаг открывает ещё больше непонятного. Мы можем только приближаться, ошибаться, проверять и снова ошибаться. Знание всегда неполное, и именно в этой неполноте мы продолжаем двигаться».
Поэтому приравнивать их — это ошибка. Одна построена так, чтобы спрятать реальность, которая не укладывается в готовый ответ.
Другая построена так, чтобы сталкиваться с этой реальностью снова и снова, даже если от этого становится некомфортно.
Называя меня сатанистом, ты на самом деле говоришь: „Тот, кто не хочет прятаться за готовым смыслом и предпочитает сталкиваться с реальностью как она есть — для тебя уже дьявол“.
Это честно показывает, что для тебя важнее утешение, чем истина.
Я же считаю наоборот: лучше жить с открытыми глазами, даже если иногда холодно и страшно, чем в тёплом, но выдуманном мире.»-
Плоскую землю придумали в 19ом веке, в какой-то американской академии.
и квантовая механика — две равноценные секты. На деле они устроены совершенно по-разному.
1. Я не говорил про квантовую механику. Это дисциплина, а я говорил про учёных, что их сектантизм в той же мере сектантизм, что и религиозный, гомеопатийный, вегетарианский, зелёный и т.п.
2. Как раз устроены сектанты строго одинаково, с точки зрения корневого когнитивного расстройства. Подмена рассудка эмоциями, фиксация на какой-то сверхидее.
Кто там из них лучше или хуже зависит от выбранной системы ценностей, та же научная секта плотно связана со скопничеством и ведёт к вымиранию популяции, поэтому если смотришь с точки зрения биологии, то она очевидным образом вредная.
Обратите внимание, я не говорю про науку, я говорю про рядовых учёных, которые очевидным образом не особо придерживаются научного принципа.
В рамках секты учёных учат двоемыслию, когда ты обязан придерживаться этого принципа в своей прикладной области и не имеешь права придерживаться в тех, на которые укажут лидеры секты.
Быть учёным в системе, но вне секты очень сложно, тут Дробышевский выгодно выделяется.Называя меня сатанистом, ты на самом деле говоришь
Ты гнойный расист, ультрахамло (кидаться на женщин это дно) и шизик, учитывая твои отношения с родителями. Самое что ни есть дно Элементов.Я же считаю наоборот: лучше жить с открытыми глазами
Тебе до этого далеко, сильно зашоренный.-
Так ты единственный "несектант" на всей земле? Это даже не позиция, а диагноз. Ты в самой классической и самой жалкой секте на свете — секте «Я один просветлённый». Таких как ты всегда полно: от мелких гуру до великих вождей. Так что давай уже без цирка: либо ты признаёшь, что сам сидишь по уши в своей уютной секте «я выше всех вас, глупых сектантов», либо начинаешь принимать таблетки от мании величия.
-
-
-
-
-
а вынуждены именно что противостоять самой идее самореплицирующихся рибозимов в той или иной форме
Ну зачем приписывать оппонентам то, что они "не исповедуют"? Вопрос, который ставят креационисты (по крайней мере сейчас, не знаю как было ранее), не в невозможности саморепликации первой формы жизни, а в вероятности этого процесса. Самореплицироваться и целый человек может, только добавьте достаточно мультивселенных)).
Есть некий порог, после которого вероятность случайности какого-то события выглядит менее убедительно, чем вероятность сознательного процесса. Соответственно, чем выше порог невероятности самосборки первой формы жизни, тем выше вероятность ее искусственного создания (или наличия неприлично большой Вселенной).-
Скажем, конкретно я думаю, что жизнь именно что самозародилась на Земле, что её появление и развитие до человека является экстремально низковероятным событием, а с изобретением ИИ человечество прекращается и постцивилизация остаётся точкой в пространстве. Поэтому мы не видим других (иначе бы вся галактика была забита просто из-за длительного времени).
Но в то же время я сам делал множество разных симуляторов, в том числе жизни, где я руками создавал стартовые конфигурации, чтобы посмотреть во что всё разовъётся. Ну удачи моим жучкам понять откуда они возникли :)
Причём фишка в том, что искусственных вселенных намного больше, чем реальных. Это как с мозгом Больцмана, чисто вероятностно мы можем ожидать того, что кто-то извне запустил вселенную в нужной конфигурации, чтобы посмотреть нынешние новости.
-
-
-
-
Когда мне кто-то говорит о том, что ему нужно больше времени чтобы самозародить жизнь, я вспоминаю Ходжу Насреддина, пообещавшего падишаху научить осла говорить за 20 лет, взяв за это приличную сумму денег))
-
История про Насреддина — это как раз про мошенничество со сроками: пообещать чудо побыстрее, взять деньги и надеяться, что за 20 лет либо осёл сдохнет, либо падишах.
А когда кто-то требует «покажите самозарождение жизни прямо сейчас», он, по сути, мыслит ровно так же — только наоборот: хочет невозможного в заведомо недостаточный срок, а потом делает вид, что раз не получилось «за 20 лет», значит, и за миллиарды невозможно.
Наука в этом месте выглядит более серьезно: она не торгуется за удобные сроки, а честно говорит — у нас есть миллиарды лет, и за это время из одноклеточных появился человек.
Так что если уж искать Насреддина, то он не на стороне науки, а на стороне тех, кто требует, чтобы осёл заговорил прямо сейчас — или объявляет, что это в принципе невозможно.
-
-
Тут в основном кипятятся сектанты-сторонники зарождения. Бездумно орут против: Kordum, за: Вячеслав Рогожин, kbob и aureliano. Логически правильно сказать, что кипятятся сторонники зарождения.
У них есть альтернативная теория?
А её не нужно, чтобы критиковать эту :) Это у вас ложная дихотомия.Теория самозарождения жизни на земле вполне опровергаема
Скорее всего нет.стоит только на Марсе или каком-нибудь астероиде найти РНК с земными нуклеотидами или белки из земных аминокислот
Это ни на что не ответит. И тут речь не про Землю, а про самозарождение вообще. Предположение, что она зародилась на Марсе, на мой взгляд предельно нелепо, но если даже допустить, то там то она тоже должна была возникнуть в каком-то процессе. Это ни на что не отвечает. -
Фундаментальная наука, некоторым образом, ищет ключи под фонарём. Ну просто потому, что вокруг бушующий океан, где ты точно ничего не найдёшь, хотя ключи могут быть именно там. Но под фонарём их хотя бы возможно найти, и проверить там быстрее всего.
Повторюсь, для меня концепция самозарождения жизни кажется самой осмысленной, но исключать того, что оно было сконфигурировано извне тоже нельзя, но это непроверяемо и даже непознаваемо, поэтому в ту сторону нет особого смысла копать. Хотя есть предпосылки в виде самосознания и разума, которые кажется не особо физичны, которые заставляют на эту тему хотя бы просто говорить.
Можно представить себе такую картину: мы делаем симуляторы, физические, биологические, игры. В них может жить жизнь (для нас она искусственная, но для неё самая что ни есть настоящая). Для нас эти симуляторы являются простыми, там сильно упрощённые законы, объём и прочее.
Но можно сделать некоторое преобразование ИЗ симулятора в нашу жизнь. Оценить рост сложности и объёма. А потом применить это преобразование уже к нашей жизни, чтобы хотя бы примерно оценить уровень сложности с которым мы можем иметь дело.
Сейчас учёные кидаются словом мультивселенная, но смысл этой теории не выше, а может и ниже, чем у предположения, что мы в симуляторе. Мультивселенных мы вообще не наблюдаем, а вот симуляторов море.
Я не к тому, что не надо копать, копать надо. Просто нужно не забывать, что альтернативы возможны и возможно можно подобрать эксперименты, очень сложные и нонкормистские, которые смогут получить какие-то свидетельства вне мейнстрима.-
Ещё в тему мультивселенных. Вот есть модель мозга Больцмана, которая говорит, что во вселенной вероятность самозарождения разума из вакуума во вселенной может быть многократно выше, чем зарождение биологическим путём. Типа если вселенная бесконечна, то случайно взятый интеллект скорее всего будет самозарождённым из вакуума.
Отсюда сразу вопрос, если говорить про мозг Больцмана, то какой смысл вообще говорить про нашу физику, может нас глючит, что физика такая, какой мы её видим, а может мозг Больцмана зарождается в другой физике, где это попроще, просто каналы восприятия видят именно нашу.
Поэтому приплетая мультивселенную к вопросу появления жизни, нужно понимать, что это возможно бессмысленно. С тем же успехом в мультивселенной и мозг Больцмана появится, или появится жизни, которая создаст компьютеры, в которых хаос в битах создаст симулятор в котором мы живём. Смысл одинаковый, никакой.-
Вот есть модель мозга Больцмана, которая говорит, что во вселенной вероятность самозарождения разума из вакуума во вселенной может быть многократно выше, чем зарождение биологическим путём. Типа если вселенная бесконечна, то случайно взятый интеллект скорее всего будет самозарождённым из вакуума.
Да не факт вообще. И даже не в этом дело. ОК - пусть есть где-то сферический-в-вакууме самозарождённый "мозг". С "глазами". То есть "разумный", "сознательный" наблюдатель. Мозг и наблюдатель существуют не просто так - они что-то наблюдают и обрабатывают. Вопрос: ЧТО этот самый БМ будет наблюдать и главное - КАК ДОЛГО. Концепция БМ вообще не занимается рассмотрением вопроса о том, что будет ПОСЛЕ того, как он возник. А, согласно всё той же термодинамике, этот самый только что возникший "мозг" с высочайшей вероятностью распадётся в следующее же мгновение. Ещё больше - в следующую секунду, ещё больше в следующую минуту. И какое тогда значение имеет - был ли он где-то вообще?
Тогда как только результаты работы дарвинвского механизма ИЗНАЧАЛЬНО, просто по определению, должны получаться и получаются от раза к разу всё более и более УСТОЙЧИВЫМИ в среде своего появления:
https://elementy.ru/nauchno-populyarnaya_biblioteka/430413/Evolyutsiya_kak_soprotivlenie_entropii
Так что разумный наблюдатель как таковой во вселенной получается со сколь нибудь существенной вероятностью только в результате работы дарвиновского механизма.-
Да не факт вообще.
В таких масштабах познаваемости вообще всё не факт. Это утверждение требует, чтобы вселенная была околобесконечна в будущем времени, что выходит за пределы нашего познания. Туда же в той же мере относится мультивселенная, теория симулятора (которая включает в себя прокариотный креационизм). Собственно сама концепция Мозга Больцмана была придумана, чтобы оценить рамки возможного познания.
Как итог, упоминание мультивселенной в статье имеет не больше смысла, чем бога, и меньше, чем Энгельса.С "глазами". То есть "разумный", "сознательный" наблюдатель.
Вы здесь смешиваете три очень разных сущности.
1. Глаза. Это некий канал восприятия информации, с точки зрения данного обсуждения в них может подаваться и некая предзаписанная или генеративная информация. Очевидным образом может образоваться двойной мозг, где одна половина генерирует картинку мира, а другая на неё смотрит.
2. Сам мозг и длительность его существования. Вы же не знаете, сколько сами существуете. У вас есть память, но вот я сейчас работаю со сложными вычислениями, и чтобы отлаживать новую фазу, я данные предыдущей фазы сохраняю и их уже не запускаю. Новой фазе кажется, что она непрерывна, в то время, как я много раз запускаю её из среза.
3. Для меня непонятен эффект самосознания, который кажется нефизичным, какой-то проекцией без понятных свойств. Т.е. мозг может функционировать автономно, примерно, как та же нейронка (на другом формате математики, но похожем принципе). Но в какие-то моменты времени возникает сознания, которая анализирует, что там нагенерировал мозг и принимает решения. И вроде бы она не нужна, но ощущается, поэтому обсуждается. Это полностью аналогично действию, когда игроки в компьютерных играх ассоциируют себя с персонажем игры, который прекрасно может существовать и без разумного вмешательства, но оно всё-равно возникает и в этот момент виртуальный персонаж деятельно ощущает себя реальным.высочайшей вероятностью распадётся в следующее же мгновение
Мы говорим про мысленный эксперимент, в рамках которого мы в рамках физической возможности можем предположить любую интересную нам конфигурацию в пространстве и времени.
Вроде как физика их позволяет, а значит, что если вселенная бесконечна, то случайный разум в ней будет вакуумным, что можно принять за разумное предположение.механизма ИЗНАЧАЛЬНО
Злоупотребление выделением текста является признаком некоторого расстройства.Так что разумный наблюдатель как таковой во вселенной получается со сколь нибудь существенной вероятностью только в результате работы дарвиновского механизма.
В рамках бесконечной системы и устойчивого вакуума - нет, почти любой разумный наблюдатель появляется не из эволюции-
В таких масштабах познаваемости вообще всё не факт.
Тогда и твои умозаключения тоже не факт - зачем тогда ты о чем-то рассуждаешь "в таких масштабах"? Так что давай уже без цирка: либо ты признаёшь, что сам сидишь по уши в своей уютной секте «я выше всех вас, глупых сектантов, потому что ничему не верю»», либо начинаешь принимать таблетки от мании величия.
Не стоит уходить в такие масштабы где только твои умозаключения являются фактом для тебя... -
Вы же не знаете, сколько сами существуете. У вас есть память
Не только. Я могу непосредственно наблюдать следы старения организма даже внешние. Для данной задачи этого достаточно. Это прямое следствие работы Второго начала термодинамики, которое одно только и определяет направленность стрелы времени и по статистике которой вычисляется в том числе и концепция БМ. Фокус только в том, что после оного БМ появления, частицы, его составляющие, двигаться не перестают. А причин, заставляющих их сохранять случайно получившуюся конфигурацию - нет. Поэтому она распадается в следующее мгновение. Вот и весь ВАш БМ.Мы говорим про мысленный эксперимент, в рамках которого мы в рамках физической возможности можем предположить любую интересную нам конфигурацию в пространстве и времени.
Конфигурация, в которой случайный разум тут же распадается не успев обработать и пару своих мыслей может быть кому-то и интересен, но точно не мне. А всем моим знакомым коллегам - тем более.
Вроде как физика их позволяет, а значит, что если вселенная бесконечна, то случайный разум в ней будет вакуумным, что можно принять за разумное предположение.
Вы и дальше можете измышлять любые умозрительные, но совершенно бесполезные конструкции - но уже самостоятельно. Рассуждения о среднем количестве ангелов или демонов Максвелла, умещающемся на кончике иглы мне не интересны совершенно.Злоупотребление выделением текста является признаком некоторого расстройства.
Тогда все школьные учителя и лекторы - больные люди?В рамках бесконечной системы
Предмет естественных наук - только конечные системы.
Актуальные бесконечности - предмет лишь математики.-
Я могу непосредственно наблюдать следы старения организма
А с каплей крови в мозгу, в неправильном месте, можете наблюдать много чего ещё. Вы, кажется, не очень в абстрактное мышление умеете.Это прямое следствие работы Второго начала термодинамики
Вы его не понимаете, если что. Оно 1. носит вероятностный характер и допускает всякое, 2. оно тут неприменимо.Поэтому она распадается в следующее мгновение
Не имеет значения, тем более, что ничего не мешает ему появиться в скафандре. Ещё раз про эмоции и связанную с этим неспособность в сложный анализ.пару своих мыслей может быть кому-то и интересен, но точно не мне
Ну и не встревали бы в разговор.но совершенно бесполезные конструкции
Ну свинье в хлеву и компьютеры бесполезны, чего лезете-то?Тогда все школьные учителя и лекторы - больные люди?
Они такого не делают, кроме как по специально предназначенного для этого случаям. Вы не в школе. И вы да, определённо страдаете каким-то психозом. Но вроде мы его уже обсуждали.Предмет естественных наук - только конечные системы.
Вам то откуда знать про науки.-
А с каплей крови в мозгу, в неправильном месте, можете наблюдать много чего ещё.
"Если-бы-да-кабы-да-во-рту-росли-грибы...." Вы, кажется, не очень в абстрактное мышление умеете.
Да - я не очень люблю пустые фантазии, не имеющие отношения к реальности, которые вы почему-то называете "абстрактным мышлением".Оно 1. носит вероятностный характер
Как и ваши модели БМ. И?и допускает всякое
но не перестаёт действовать и после этого.2. оно тут неприменимо.
Потому что Вы так решили?Не имеет значения
Тогда для меня не имеют заначения все те невидимые розовые единороги, которых вы тут сочиняете. Потому дальше можете обсуждать их сами с собой.Не имеет значения, тем более, что ничего не мешает ему появиться в скафандре.
Ещё минус порядки вероятности. Какого объёма/массы "скафандр"? Вы уверены что вероятность такой конструкции в итоге должна оказаться существенно меньше вероятности "случайной сборки" обводнённой скалистой планеты у жёлтой звезды главной последовательности?Ещё раз про эмоции и связанную с этим неспособность в сложный анализ.
Увиденные ВАми в зеркале?Ну и не встревали бы в разговор.
Просто прямо уведомляю. А встревать или не встревать в публчный разговор буду исключительно по собственным соображениям.Ну свинье в хлеву и компьютеры бесполезны, чего лезете-то?
Некорректная аналогия. Компьютеры - наблюдаемы. БМ - нет.И вы да, определённо страдаете каким-то психозом.
Можно увидеть ВАш врачебный диплом и сертификат по психиатрии, дабы все читатели могли убедиться, что Вы обладаете квалификацией в постановке психиатрических диагнозов?Вам то откуда знать про науки.
Может быть потому, что обладаю учёной степенью?-
"Если-бы-да-кабы-да-во-рту-росли-грибы....&
У вас не хватает интеллектуальных способностей, чтобы вести разговор на эту тему, понимаете? Ваш психоз вас ограничивает. Я не просто так указывал на ваше увлечение выделениями текста, если бы мне кто-то указал на такие объективные проявления расстройства то я бы напрягся.quot; Да - я не очень люблю пустые фантазии
Ну как свинья про компьютеры говорит. Компьютеры тоже пустые фантазии для тех, кто не способен ходить по всему пространству понятий.но не перестаёт действовать и после этого.
Оно не перестаёт, только вы не понимаете, что это значит. Для вас границы применимости термодинамики это "пустые фантазии".Ещё минус порядки вероятности.
Нет тут никакого минус порядка.Просто прямо уведомляю
Вы только свой диагноз описываете, но мы с вами уже это разбирали в прошлый раз, поэтому ничего особо нового не будет. Обратите внимание, это конкретно ваш диагноз.БМ - нет
Ещё как наблюдаемы. Динамический эффект Казимира тот же самый. Остальное всё вопрос вероятности, т.е. как сказано изначально - упирается в предполагаемую длительность жизни вселенной с подходящим вакуумом.диплом и сертификат по психиатрии
Я вам указывал и указываю на конкретные проблемы. Из ваших собственных текстов. Вы же сами указываете пределы своих аналитических способностей. Пока нет нарушения изложенной мною теории, она обоснована исторически и с двух сторон в данной теме.Может быть потому, что обладаю учёной степенью?
Где связь?-
У вас не хватает интеллектуальных способностей, чтобы вести разговор на эту тему, понимаете? Ваш психоз вас ограничивает.
Я бы всё же рекомендовал ВАм убирать зеркало от монитора, когда вы что-то отвечаете другим в интернете.Компьютеры тоже пустые фантазии для тех, кто не способен ходить по всему пространству понятий.
Почему аналогия умозрительного БМ с компьютерами некорректна - вам уже писалось: компьютеры наблюдаемы, а БМ - нет. Более того - для БМ даже не существует чёткого определения какому диапазону физических свойств должен удовлетворять его носитель.Ещё как наблюдаемы. Динамический эффект Казимира тот же самый.
И какова же связь между умозрительной концепцией больцмановского мозга и оным эффектом?Оно не перестаёт, только вы не понимаете, что это значит.
Прекрасно понимаю и прямо написал что: разрушает случайно собранную систему частиц.Нет тут никакого минус порядка.
Есть просто элементарно в результате увеличения минимального размера (объёма, числа задействованных частиц) требуемой системы БМ+скафандр > простоБМ в вакууме.Я вам указывал и указываю на конкретные проблемы.
Это не проблемы, а особенности стиля, которые были убраны сразу, как на них указал собеседник. Не нужны выделения шрифтом - ОК. Сверхценным значением (в отличие как раз от тех, у кого и правда могут наблюдаться шизотипические явления) для меня они не обладают.Где связь?
Связь между обладанием учёной степенью и знанием о науках - и правда - где же? Ну очень сложно её найти, да?-
Я бы всё же рекомендовал ВАм убирать зеркало от монитора, когда вы что-то отвечаете другим в интернете.
На эту тему бы я подумал, если бы не ваши демонстративно ограниченные рассуждения о природе скафандра, которые показывают узость взглядов.
Я вот ною от нашего общества, что узость взглядов возводится в абсолют, в том числе среди учёных, которые как бы должны были бы уметь применять научный принцип, но не просто не умеют, а кичатся этим самым неумением. Отсюда у страны много проблем.аналогия умозрительного БМ с компьютерами некорректна
БМ не с потолка взяли, это следствие найденных физических законов. То что вы их не видите, это ваши проблемы, а не преимущество.больцмановского мозга и оным эффектом?
Прямая, БМ возникает как часть этого эффекта.разрушает случайно собранную систему частиц.
Емнип, БМ изначально был придуман для химических систем, где летают конкретные атомы. Позже его перенесли на квантовый мир, где существующих атомов не нужно. Мне кажется, что вы имеете в виду химический вариант. Для химической термодинамике во всех учебниках пишут специальную оговорку, что второе начало не выполняется на космологических масштабах.Это не проблемы, а особенности стиля
Это признак эмоциональности, эмоциональность в интеллектуальной среде означает снижение аналитических способностей, поэтому рекомендуют не злоупотреблять.Связь между обладанием учёной степенью и знанием о науках - и правда - где же? Ну очень сложно её найти, да?
Связи тут нет. У нас не учат думать (а это настолько же необходимо, как учиться контролировать тело для спорта). Кроме того в научной среде процветает сектантизм, который прямо запрещает думать по определённым темам, иначе ты сразу теряешь авторитет в среде. Приходится вырабатывать странную форму двоемыслия, зачастую не до конца осознанную, потому, что думать и анализировать не учат. Это приводит к выбросам, приводящим к академикам-гомеопатам, фоменковщине, антивоенным подписантам и прочей ереси.
Поэтому от человека со степенью можно ожидать, что у него есть бумажка, и с определённой натяжкой, что он разбирается в своей теме. Но нельзя ожидать, что он умный (умеет находить лазейки, работать с невидимым), умеет применять научный принцип (ко всем аспектам познания мира), или понимает особенности нашего интеллекта (когнитивные искажения). Всему этому нужно учиться отдельно, и если бы вы это делали, то у вас и вопросов про научную степень не возникло.-
БМ не с потолка взяли, это следствие найденных физических законов.
У физических законов может быть множество "следствий", к наблюдаемой физической реальности отношения не имеющих.
Чрезмерное увлечение такими следствиями подобно этой картине: https://xkcd.ru/605/Прямая, БМ возникает как часть этого эффекта.
Каким образом? С какой вероятностью? Что физически должен представлять из себя такой БМ? Есть публкация на эту тему?Позже его перенесли на квантовый мир, где существующих атомов не нужно.
То есть, проще гоовря, в "виртуальную реальность". Ну, там и улыбки без кота и невидимые розовые единороги могут быть. Но более настоящими _тут_, в нашей макрореальности они не становятся.Мне кажется, что вы имеете в виду химический вариант.
В смысле состоящий из множества стабильных частиц при температурах выше лямбда-точки гелия. Вообще-то это и есть та область мира, для описания которой и открыта статфизика-термодинамика. Тем же Л. Больцманом.Для химической термодинамике во всех учебниках пишут специальную оговорку, что второе начало не выполняется на космологических масштабах.
Космологические масштабы - они разные бывают кагбе.Это признак эмоциональности, эмоциональность в интеллектуальной среде означает снижение аналитических способностей, поэтому рекомендуют не злоупотреблять.
В данном случае - это лишь акцентирование внимания на важных по мнению докладчика местах для читателя/слушателя с меньшим или неизвестным уровнем подготовки.антивоенным подписантам
Какое отношение политическая позиция какой-либо части любого проф.-сообщества имеет к всему вышевамиперечисленному? Учёный не должен иметь права высказать своё мнение по тому или иному политическому вопросу (в т.ч. и коллективно)?Я вот ною от нашего общества, что узость взглядов возводится в абсолют
А я вот не ною, потому что с детства приучен, что быть нытиком - это позорно. И нытиков сам презираю. Чем далее - тем более.узость взглядов возводится в абсолют, в том числе среди учёных, которые как бы должны были бы уметь применять научный принцип, но не просто не умеют, а кичатся этим самым неумением.
Нет никакого одного "научного принципа" есть научный _метод_, включающий в себя по меньшей мере четыре принципа, по которым должна строиться любая научная гипотеза:
1. Связность И непротиворечивость;
2. Конкретность описываемых явлений (конечность, ограниченность);
3. Проверяемость (повторяемость);
4. Минимальность (неизбыточность).
У умозрительных концепций типа БМ или струнных моделей с проверяемостью в реальных наблюдениях дело обстоит совершенно никак.-
Чрезмерное увлечение такими следствиями подобно этой
Не тот случай. БМ это не про материю с отрицательной энергией, а вполне конкретная волновая функция подтверждённая в экспериментах. К тому же это в общем-то мысленный эксперимент призванный конкретизировать наши ограничения и воспринимать его нужно именно в этом ключе.
Понимание свои ограничений это ключевая особенность умного человека (мало понимать, что ты глупый и постоянно ошибающийся, нужно ещё понимать контуры этой глупости и ценность ошибок).С какой вероятностью?
Из того, что я видел, в видимой вселенной должен возникать за 10^10^50 лет. Это к вопросу из предыдущего абзаца, где вы бросились говорить на тему не удосужившись ознакомиться с предметом разговора и не попытавшись понять рамки применимости.Но более настоящими _тут_, в нашей макрореальности они не становятся
Макрореальность это просто слова, а реально на таких масштабах мы находится в нефальсифицируемых теориях, и говорить настоящая наша реальность, выдумка или виртуальная - не можем.
Из самой концепции, что наша реальность макро, например следует, что она в принципе непознаваема, иначе в силу бритвы оккама нужно будет признать, что мы просто завихрение в формуле.статфизика-термодинамика. Тем же Л. Больцманом.
Ну да, давайте ещё древних греков вспомним.это лишь акцентирование внимания на важных по мнению докладчика
Ну да, читатели же тупые, НуЖНо ОбЯЗАтельНО ВыклЮЧИТЬ иМ КогнИТИвочКу!11
Как я сказал, вы демонстрируете эмоциональность, которая подавляет аналитику, это вредно прежде всего для себя самого.Какое отношение политическая позиция какой-либо части любого проф.-сообщества имеет к всему
Знал, что зацепитесь :) Есть набор маркеров, которые железно работают по сектантам (разные маркеры для разных областей). Есть довольно ужасные, которые я не озвучиваю из уважения конкретно к Элементам, но работающие с практически 100% (выше 99).
Это не политическая позиция, это секта, в самом прямом смысле этого слова. У сектантов табуирован анализ в этой области. Самый грубый отказ от применения научной парадигмы в области, на которую запрещено обращать внимание в рамках корпорации.
Можно даже научно обоснованно сказать, любой, кто против СВО - психопат (с элементами шизофрении или социопатии) нуждающийся в квалифицированном лечении (неспособный преодолеть психический блок самостоятельно), который вредит себе и соседям, но я не буду развивать эту тему, вы не поймёте в силу ранее описанных причин.
Ещё одно из проявлений секты это скопничество. Если эту тему поразвивать, то биологи начинают яростно отрицать теорию эволюции :)
Ну и задачка про кислород для дыхания, тут обычно отрицают закон сохранения.
Поэтому я предпочитаю объективизировать свои концепции через привязки к объективным фактам и считать когнитивные искажения приматом над моими мыслями.
Если тысяча человек мне доказывает, что дерево может выделить кислорода в несколько раз больше своего веса, то эти 1000 человек неправы. Если человек считает, что изнасилования детей на глазах у матери приемлемы, то его нужно ликвидировать. Реальность просто нужно признать как истину, невзирая на социальное давление, иначе становишься сектантом, что влечёт за собой другие неприятные последствия.А я вот не ною, потому что с детства приучен, что быть нытиком - это позорно.
Но и не думаете, причём ещё и бравируете этим, а на мой взгляд это более позорно. Всему своё место и время.Нет никакого одного "научного принципа"
Это вы построение теории описываете, это уже следствие, инструмент.
Основа основ идея, что мы можем и скорее всего заблуждаемся, что наша животная природа постоянно пытается нас обмануть и упростить. Признавая факт того, что мы в основном ошибаемся (чего конкретно вы не признаёте), можно выработать набор формальных правил обеспечивающих поиск наших ошибок. Не подтверждения идей, а именно ошибок. И умный человек радуется тому, что находит свою ошибку и ничего не испытывает в случае подтверждения своих взглядов (нулевая ценность). Без этого понимания всё остальное научное познание не имеет смысла. Сектанты фокусируются на какой-то сверхидее, подавляя эмоциями возможность осознать ошибку.-
БМ это не про материю с отрицательной энергией, а вполне конкретная волновая функция подтверждённая в экспериментах.
А можно ссылку на публикацию, где это описывается?К тому же это в общем-то мысленный эксперимент призванный конкретизировать наши ограничения и воспринимать его нужно именно в этом ключе.
Вот его продолжение в виде тут же следующего разрушения БМ после его формирования - и есть ограничение данной модели.а реально на таких масштабах мы находится в нефальсифицируемых теориях
То есть ненаучных по определению.и говорить настоящая наша реальность, выдумка или виртуальная - не можем.
Значит - как и делать какие-либо иные содержательные утверждения.Из самой концепции, что наша реальность макро, например следует, что она в принципе непознаваема, иначе в силу бритвы оккама нужно будет признать, что мы просто завихрение в формуле.
Это ничего не меняет в действительно значимой сути. Ни у какой науки давным-давно не стоит цели познать что-то абсолютно. Даже у математики после Гёделя. Наука - это ИНСТРУМЕНТ познания. У любого инструмента есть границы применимости.Ну да, читатели же тупые,
Заметте - не я это написал. ;~]Как я сказал, вы демонстрируете эмоциональность
"Вы считаете что читатели тупые...!", "Я просто ною от...!" - это конечно, строго академическая речь, ага - а демонстрирует эмоциональность оппонент, угу.
Вижу, что добрым советом убрать зеркало от монитора Вы так и не воспользовались. Жаль.Это не политическая позиция, это секта, в самом прямом смысле этого слова.
Потому что вы такрешли? ответ на свой вопрос я так и не услышал.
Вы отказываете какому-либо профессиональному сообществу (в данном случае научному) - да или даже любому общественному объединению (хоть секте) - публично коллективно выражать свою позицию по какому-либо политическому вопросу?
Вы считаете, что те объединения, что признаны "сектами" каким-либо экспертом (например Вами) следует в обязательном порядке уничтожать?Если человек считает, что изнасилования детей на глазах у матери приемлемы, то его нужно ликвидировать.
Публичным четвертованием? Или лучше сожжением заживо?Если тысяча человек мне доказывает, что дерево может выделить кислорода в несколько раз больше своего веса, то эти 1000 человек неправы.
Их тоже нужно за это карать?-
Я почитал, как вы рассказываете, что секты это хорошо, быть тупым законное и почётное право, насиловать детей нормально и решил, что больше не буду пытаться вам указывать на то, как можно было бы вылечиться самостоятельно. Вам нужно в тюрьму или дурку.
-
^- вот, что происходит, когда не следуешь доброму совету убрать зеркало от монитора при написании ответа в интернете.
-
Ты самый каноничный фашист, который многократно себя самого описал, восхваляя тупость, сектантство и считая, что самые страшные преступления из возможных приемлемы (это собственно является разделяющей чертой между сторонниками России и Запада. Сторонник России сказал бы, что это неприемлемо, чего никогда не скажет сторонник Запада - это ещё один важный детектор).
Это не зеркало, это ты сам про себя рассказал в деталях рассказал.
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
"Вот есть модель мозга Больцмана, которая говорит, что во вселенной вероятность самозарождения разума из вакуума во вселенной может быть многократно выше, чем зарождение биологическим путём."
Вот именно, что может быть, а может и не быть. Для того, чтобы такое могло быть, помимо мультивселенной, состоящей из бесконечного или почти бесконечного числа вселенных, необходима ещё и безумная скорость их образования. Действительно, если число вселенных удваивается каждую секунду или наносекунду, то в каждый момент времени вселенных, в которых жизнь могла успеть возникнуть и проэволюционировать до наших мозгов, будет на много порядков меньше, чем молодых вселенных, которым не хватило времени для эволюции, но в некоторых из которых могли случайно возникнуть такие мозги. Однако измените всего одно допущение: в бесконечной мультивселенной число вселенных удваивается не каждую наносекунду, а каждые 10 млрд лет, и плотность вероятности появления больцмановских мозгов будет на много порядков меньше плотности вероятности дарвиновской эволюции. Т. е. в случае с мозгами больцмана делается не одно, а сразу несколько совершенно произвольных недоказуемых предположений. В этой же статье мультивселенная упоминается, как плохой способ объяснения чего либо, включая абиогенез, и показывается, что можно обойтись и без неё.-
Речь про нашу вселенную живущую околобесконечное время. Мультивселенная это про другое. Её зря упоминают, а указанная работа ничего особо не показывает.
В рибозимах есть высоко консервативные участки состоящие из десятка нуклеотидов образующих ядро рибосомы. Вот если бы нашли молекулу с соответствующей активностью похожую на эти центры - это было бы прикольней.-
Так в вечной или просто очень долго живущей вселенной плотность вероятности дарвиновской эволюции намного больше, чем спонтанного появления больцмановских мозгов. Настолько же порядков больше, насколько меньше показатель степени 45 показателя степени числа молекул и других частиц в мозге.
-
Из-за расширения вселенной и исчерпания топлива - вся видимая нами эволюция закончится за малое время в общем масштабе. Ну и вы зря цепляетесь к этим 45, они ничего не значат. Маловероятно, что жизнь стартовала бы без доступа к залежам того же ZnS, который выпал на Землю после образования Луны и постоянно парящих геотермальных источников, что происходит краткий миг в эволюции планеты.
-
"Из-за расширения вселенной и исчерпания топлива - вся видимая нами эволюция закончится за малое время в общем масштабе."
Об этом я не думал. Интересная мысль. И даже расширение не нужно — достаточно тепловой смерти статической вселенной. Но, думаю, в этом случае вакуумная флуктуация, которая сможет породить новый большой взрыв, более вероятна, чем спонтанное появление больцмановского мозга из вакуума.
"Маловероятно, что жизнь стартовала бы без доступа к залежам того же ZnS, который выпал на Землю после образования Луны и постоянно парящих геотермальных источников"
Так я же не отрицаю необходимости внешних условий. В эксперименте тоже создали условия для роста РНК. Она же не сама собой у них появилась. Но вы хотите, чтобы условия в лаборатории были точно такими же, как на древней Земле. Тогда дайте и им как минимум несколько миллионов лет, которые были у древней Земли.-
Не нужно делать "такие же" условия, нужно делать правдоподобные. Для примера приведу допустим на земле было N точек зарождения, по гауссову распределению, из них в 5% окажутся условия плохие и медленная эволюция на те самые миллионы лет, в 15% хреновенькие на сотни тысяч лет, в 60% средние на десятки тысяч лет, в 15% хорошие на тысячи лет и в 5% суперские на сотни лет. Задача ученого найти те самые последние условия, привести их к еще более выгодному формату для лаборатории и поставить эксперимент, тогда с большой долей вероятности сроки сократятся до десятков или менее 10 лет. Что нередко в лабораторных исследованиях. Я сам лично мембраны гонял 3 года исследуя их старение.
-
Не нужно делать "такие же" условия, нужно делать правдоподобные.
Мне кажется, что это будет уже другим экспериментом. В данном опыте они подобрали более-менее жизнеспособную короткую РНК. Причём сделали это не случайным перебором, а используя инженерию, т. к. миллионов лет у них нет. В другом можно проверить, возможно ли за разумное время собрать РНК такой же длины в похожих на древнюю Землю условиях чисто случайно. Но требовать, чтоб такая РНК была бы ещё и жизнеспособной с 1-ого или 100501-ого раза — перебор. Если можно собрать случайно какую-нибудь последовательность такой длины, а из всех таких последовательностей ~10^11 являются более-менее рабочими, то неизбежность абиогенеза в этих условиях можно считать доказанной. Правда, желательно ещё доработать 45-нт ДНК, чтобы она эволюционировала быстрее, чем вырождалась или разрушалась. Но это всё разные эксперименты. Объединить их можно, только если есть миллионы лет. И главный из этих 3 экспериментов — 1-й, т. е. создание короткой рабочей последовательности, что они и сделали. Или я чего-то не понимаю?-
Ну не, про жизнеспособность тут речи и близко не идёт.
Это как с термоядом, реакции совершать умеем, а термоядерную электростанцию не построили и скорее всего никогда не построим. Причём настоящие термоядерные реакции на Солнце идут по совсем другим принципам, чем те, что мы ищем (читай - реальное происхождение жизни происходило далеко не так, как предполагается в статье).-
Поэтому я и написал "более-менее жизнеспособная", имея в виду способность реплицироваться, и упомянул про 3-й эксперимент, связанный с поиском действительно жизнеспособной РНК примерно таких же размеров (до 50 нт., например.)
А энергетически выгодный термояд уже существует. Проблема в том, что он пока экономически нерентабелен. Но экономическая проблема не фундаментальна, т. к. связана, с одной стороны, со стоимостью технологий, а с другой — с рентабельностью альтернативных технологий, которая тоже может уменьшаться по мере исчерпания ресурсов. И принцип там одинаков: разогрев плазмы. Различие же заключается только в удержании этой плазмы в одном месте.-
Поэтому я и написал "более-менее жизнеспособная"
Я по жизни очень люблю такие упрощения.
- смотрите, вот рубль. Рубль это деньги. Деньги это бюджет. Я даю вам рубль, значит закрываю бюджет. Давайте мне премию.
Нет, это даже не рядом.А энергетически выгодный термояд уже существует.
Нет не существует и очень маловероятно, что он будет хоть в какой-то осмысленной форме.с рентабельностью альтернативных технологий
Они не рентабельны.И принцип там одинаков: разогрев плазмы.
Нет. На Земле ищут энергичное преодоление кулоновского барьера, а на солнце термояд реализуется через туннелирование. Малосвязанные между собой вещи.
Солнечный термояд с его энерговыделением меньше, чем у гнилых листьев тут никому не нужен.-
Я по жизни очень люблю такие упрощения.
Сейчас это вопрос веры. Но, думаю, что в течение нескольких ближайших лет, а может, и этого года, такая жизнеспособная РНК до 50 нт. будет создана. Ведь ещё несколько лет назад речь шла о 90-х степенях десяти."А энергетически выгодный термояд уже существует."
См. статью в Википедии "Инерциальный управляемый термоядерный синтез": "КПД установки в этом эксперименте — менее 1 %".
Нет не существует
Менее 1, но больше 0 — энергетически выгодно. Разумеется, про экономическую рентабельность речи пока нет.Солнечный термояд с его энерговыделением меньше, чем у гнилых листьев тут никому не нужен.
Не нужен и невозможен — разные вещи.-
Сейчас это вопрос веры.
Ну с верой в церковь, как говориться. Посмотрим.Инерциальный управляемый термоядерный синтез
Это имеет только военное применение, ура, нас будут больше-лучше убивать. Там энерговыделение в сотни раз меньше, чем затраченное. Когда говорят про положительные значения берут одну стадию в сферовакумных условиях игнорируя огромный объём необходимой и затратной инфраструктуры.Не нужен и невозможен — разные вещи.
Это тут не к месту. Земной термояд не имеет ничего общего с солнечным и шансов на его полезную реализацию пока не просматривается даже в импульсной форме ракетных двигателей.-
Когда говорят про положительные значения берут одну стадию в сферовакумных условиях
Согласен. Сейчас посмотрел эту статью внимательней, — там написана чушь про 1%. Но КПД с каждым годом удваивается. Уже есть намного лучшие данные 25 года.“Не нужен и невозможен — разные вещи.”
Вы опять про пользу. Реализовать туннелирование намного проще, потому что требует меньше энергии. И я уверен, что это много раз делали. А насколько оно может быть полезным практически, — совершенно другой вопрос.
Земной термояд не имеет ничего общего с солнечным и шансов на его полезную реализацию
-
-
-
-
-
-
-
Вы допускаете логическую ошибку, не нужна самореплицирующаяся РНК, потому что не существовало никаких условий позволяющей ей собираться, поэтому это чистый сферический конь в вакууме. А нужна РНК поддерживающая циклы гликогенеза и Креббса.
-
Но эти циклы про обмен веществ и энергию. И без наследственности всё это работать не будет. А если и будет, то какой в этом смысл, когда нет наследственности?
-
Огонь горит, Дождь идет, Ветер дует. В этом смысл.
А какой смысл в наследственности без обмена веществ и энергии?
Абиогенез и возможна жизнь дитя радиоактивности. Опыты по зарождению жизни надо ставить не в уютных лабораториях, а в ядерных реакторах.-
Ну-ну, не, это точно не так. Жизнь и радиация это настолько антагонисты, что тут нечего особо обсуждать.
-
тут нечего особо обсуждать
Рад, что Вас заинтересовала эта тема.
Почитайте про радиолиз https://ru.wikipedia.org/wiki/Радиолиз
О том, как образуются активные соединения фосфора, молекулярный водород, и куча разной органики в углистых хондритах. О жизни в недрах земли на глубине в километры.-
Тут несовместимые порядки величин.
Радиационные процессы, которые будут производить заметные количества нужных веществ в среде - просто сварят любую нежную органику.
Нет нужды придумывать, как срастить мизерное производство от радиоактивных процессов, причём так, чтобы не развалить зародыши жизни, с вулканическими процессами, которые и так имели много всего.-
Тут несовместимые порядки величин.
Тут невообразимые порядки величин.
Мелкие бактерии в горячей минерализованной воде, где большая скорость диффузии. То есть они обедают большой объем окружающего пространства. Сами при этом являются очень мелкой целью для лучей смерти. Делятся они не раз в сутки, а раз в годы, десятилетия или даже столетия. Точно не помню.Нет нужды придумывать, как срастить мизерное производство от радиоактивных процессов,
Совершенно верно.
Придумывать ничего не надо. Надо изучать то, что у нас есть прямо под ногами на большой глубине.
(кажется сайт починили)
-
-
-
-
-
-
-
-
-
-
-
-
-
Ещё в тему мультивселенных. Вот есть модель мозга БольцманаЭто просто "разные имена бога" они не фальсифицируемы в классическом поперовском смысле.
-
Полностью согласен. И не только больцмановский мозг, но и всё, связанное с антропным принципом, нефальсифицируемо.
-
Как ни крути, а в нефальсифицирумость мы так или иначе упрёмся во всех фундаментальных направлениях науки. Теорию струн, космологию, и я очень очень надеюсь, что и интеллект.
-
"нефальсифицирумость мы так или иначе упрёмся... и я очень очень надеюсь, что и интеллект."
Наверняка когда-нибудь упрёмся, хотя не думаю, что очень скоро (космология активно развивается, теория струн, похоже, всё, но есть более перспективные теории гравитации в плане фальсифицируемости.)
Но почему вы надеетесь, что упрёмся в изучении интеллекта? Т. е., по-вашему, узнать что-то наверняка хуже, чем вечно гадать об этом?
-
-
Как ни крути, а в нефальсифицирумость мы так или иначе упрёмся во всех фундаментальных направлениях науки. Теорию струн
Уже давно упёрлась. Поэтому данная группа математических моделей и не относится к предмету естественных наук. То есть не является теорией, удовлетворяющей научному методу.космологию
Космология - не теория, а просто область знания, по сути - "предельная астрофизика", изучающая условия и силы, формирующие видимую метагалактику в целом. Из получающихся в ней теорий порою следует, что видимая метагалактика - только очень небольшая часть всей подобной вселенной (инфляционная космология). Теории эти находят наблдательное подвтерждение (параметры реликтового излучения и пр.). Вот и вся история.и я очень очень надеюсь, что и интеллект.
Зависит от того как этот самый интеллект определять.
-
-
Представьте: ученик не решил задачу по математике. Это означает что задача не имеет решения? Нет — это означает что он её пока не решил. Заварзин говорит именно это: задача абиогенеза не решена. А вы читаете это как: задача не имеет решения. Это две разные фразы. Даже школьник их различает.
Вы вроде бы "педагог", должны понимать разницу.
академик Г. ЗаварзинТут вы похоже хотели сделать отсылку к авторитету. Зря, это имеет отрицательный смысл, портит мышление вам же путём перехода в эмоциональную сферу.
самовоспроизведение системы из ДНКУже эта фраза показывает, что он не обладал достаточной компетенцией для оценки проблемы. Но сравнение его идей с миром РНК показывает направление на решение - биологическая задача последовательно декомпозируется и упрощается превращаясь в химическую.
-
И между прочим Заварзин толковый ученый точно знающий что абиогенез есть и даже выбивавший институту программу финансирования на ее изучение. Посмотреть бы результаты кстати.
-
Как можно точно знать то, что ещё не раскрыто?
Ну и по меркам развития этой области знаний он функционировал считай, что в античности.-
абиогенез - это не вопрос фактологии, это вопрос логики, он не «доказывается», а постулируется как этап самоорганизации материи
а вот какими конкретными механизмами он реализовался - это уже открытый вопрос и предмет научного поиска :3-
-
это ответ по существу, утверждать наличие абиогенетического этапа становления жизни можно без раскрытия его механики (отрицание этого постулата автоматически заменяет причинно-следственность на божий замысел) :3
-
Ты вот тут сам только что утвердил божий замысел. Сойдётся на том, что в рамках твоей логики ты знаешь, что он существует.
Кстати я - реально это знаю :))-
это не божий замысел, не вера, а постулат, как начала термодинамики
это методологический выбор и фиксация рамочной терминологии, так сказать :3-
Божий замысел, абиогенез это концепции одного порядка, до тех пор пока не найден хотя бы механизм абиогенеза. Для божьего замысла у нас вон симуляции есть, где куча народу создавало жизнь.
Если не зная механизма абиогенеза, olegov утверждает, что "знаешь, что он есть", то ты в рамках той же логики, постулируя божий замысел точно так же знаешь, что он есть. Логика.
Вот когда будет найден механизм, не сама теоретическая возможность, а более менее понятная последовательность приводящая к старту дарвиновской эволюции, тогда будет корректно говорить, что мы знаем.
Пока мы знаем, что есть химия, как она работает. Мы знаем, что эта химия чрезвычайно сложным образом собрана в нас, и даже самая примитивная живая конфигурация всё-равно выглядит невероятно сложной для самосборки, даже в том виде, котором описывает olegov. Поиск абиогенеза имеет смысл, потому, что это предметный научный поиск, у нас есть для него инструментарий и есть куда закапываться. Но в некотором роде это же поиск под фонарём, потому, что почти нет указаний на то, что мы в симуляции, и нет возможности даже с этими жалкими указаниями как-то работать.-
у креационизма есть методологический изъян - он не объясняет возникновение сложности, не объясняет возникновения бога
абиогенез - это решение этой методологической проблемы, постулирование нарастающей сложности в самоорганизующихся системах, вот и всё
(но я уверен, что ты просто троллишь, а не всерьёз ведёшь эту беседу :3)-
у креационизма есть методологический изъян - он не объясняет возникновение сложности, не объясняет возникновения бога
Любая фундаментальная научная дисциплина в конце концов неизбежно упирается в нефальсифицируемость. Та же мультивселенная упомянутая в статье это тот же самый креационизм.
Я же говорю не про то, что какая-то из идей хорошая или плохая, а то, что пока нет каких то внятных аргументов в пользу одной из них они равноправны. Это так с точки зрения формальной логики.
Поэтому нельзя сказать, что ты знаешь, что абиогенез есть, мы этого не знаем. Если уверен, что есть, значит у тебя шиза, гиперидея. Это нужно лечить.абиогенез - это решение этой методологической проблемы
Это не решение, даже не теория, а просто гипотеза.
Причём проблема в том, что это единственная гипотеза, которая хотя бы претендует на фальсифицируемость (считай поиск под фонарём).
Я не говорю, что не верую в неё, вполне в неё и верую. Только это вера, а не знание.но я уверен, что ты просто троллишь
Вам нужно логику подтягивать и учиться определять контуры своего знания. За их пределами больше, чем внутри, с этим как-то нужно работать, для того логику и придумали.-
> Я же говорю не про то, что какая-то из идей хорошая или плохая, а то, что пока нет каких то внятных аргументов в пользу одной из них они равноправны. Это так с точки зрения формальной логики.
нет, в том и дело, что абиогенез - более продуктивный базовый постулат, чем креационизм, и так оно исключительно логически :3
это не вопрос фактологии, это выбор способа описывать факты (даже ещё необнаруженные)
методологический выбор, научный
креационизм не требует объяснений (редукции), он сам становится конечным объяснением, а абиогенез - требует-
нет, в том и дело, что абиогенез - более продуктивный базовый постулат, ежели креационизм, и так оно исключительно логически :3
В смысле, что больше грантов можно стрясти? Это да.
С точки зрения научного поиска они одинаково необоснованы, мы пока не видим возможности перехода к самоупорядочиванию в абиогенезе, поэтому обладают одинаковыми правами.это выбор способа описывать факты
Это ваша вкусовщина, гиперфокус на одной идее, которая блокирует рассудочную деятельность в плане возможности найти щели за которыми может открываться что-то прикольное.креационизм не требует объяснений (редукции)
Чего бы не требует?
Повторюсь, что креационизма в нашем мире много, одних только симуляторов многие тысячи насоздавали. Вот и экстраполируй в обратную сторону и пытайся найти зацепки и из них вывести контуры гипервселенной :)
-
-
-
Любая фундаментальная научная дисциплина в конце концов неизбежно упирается в нефальсифицируемость.
Это если бы познание было бы ограничено, а оно безгранично, поэтому ничего никуда не упирается в процессе самого познание. Упирается только в твоей сегодняшней картине мира, ну твоя ограниченность и так понятна.
наука будет бесконечно отодвигать границу нефальцифицируемости.
Любое познание в конце концов неизбежно упирается в ответ "я не знаю механизма" и это про еще одно имя бога.
Древний священник не мог сказать "я не знаю" - его быстро бы лишили пайки, поэтому он говорил "бог создал", а современный ученый может сказать "я не знаю", а пайка в виде грантов остается!
-
-
-
-
Тут какое дело, нету вопроса веры, есть понимание что процесс имеет кучу шажков на каждый из которых нужно придумать свой механизм а также объяснение почему он оказался выгоднее аналогичных шажков но заводящих в тупик. К примеру до понимания роли УФ света регулярно заходили в тупик получая "белковую" массу сшитую хаотически по сути белковую смолу. Пока постулировали реакции в воде ("лужа") упирались в обращение всех реакций вспять. И так далее очень много весьма сложных и даже непреодолимых на взгляд этапов находили в итоге изящное решение. Когда от "невозможно" сразу переходили к 100%-но. Соответсвенно если ученый знает возможные этапы остается их только проверить и понять почему они а не другие. Но когда таких этапов очень много, то получается картина, когда ученый знает допустим ключевые точки а не знает промежуточные. В целом вроде как нету полного механизма но есть четкое понимание что его можно найти если потратить достаточное время и средства. Поэтому это не вера, а знание. Например не нужно знать реальный механизм горения бутана, чтобы написать реакцию его горения и знать, что бутан горит.
-
Я шажки очень люблю и на шажки очень прекрасно всё разбираю (от разработки генной терапии, до всякого другого). Но эти шажки скорее про прикладное направление, инженерию или постановку экспериментов, или готовку.
А вот в научном поиске (поиска новых неведомых структур управляющих миром, и их описание позволяющее делать предсказания) мы неизбежно, по всем направлениям, упрёмся в принципиальный предел - не будет возможности понять, что в какой среде существует очередная найденная структура. Т.е. структура есть, мы можем описать её внешнюю логику, но понять из чего она состоит нет никакой физической возможности.
Это нужно просто принимать и переставать фокусироваться на какой-то одной идее, типа мультивселенной (если ты не специалист занимающийся конкретно ей).
Так же, с точки зрения познания - сама эта граница познания тоже является конкретным объектом, с которым можно работать.
И вот тут, лично мне, интересны четыре границы:
- субквантовый мир
- космологический мир
- вопросы разума (нейронки, например, это замечательный микроскоп для изучения разума, но собственно разумом их не назвать).
- и вот зарождение жизни с её, пока, кажущейся неадекватной сложностью возникновения и развития (тех же самых эукариот), которые пока никакие натяжки (включая ваши концепции, которые я в целом разделяю, как наиболее перспективное направление поиска) не объясняют.
И вот в области всех этих границ просто замечательным микроскопом являются симуляторы, возможности которых сейчас растут невероятными темпами.
-
-
-
-
-
-
-
-
да, скорее всего «минимальный репликатор», возникший в ходе абиогенеза, это вообще не отдельная молекула, а сеть реакций со стягивающимся в эдакие «протоклеточные облака» равновесием
https://chat.deepseek.com/share/2ojw8jifrd6n9gm1ps
мы в своих клетках несём из первичного бульона не только эволюционирующего репликатора, но и эволюционирующую среду вокруг него, мы не выходцы из бульона, мы и есть этот бульон, немношк гранулировавшийся и слепившийся (и научившийся поджирать что-то снаружи этой лужи и выводя из этой лужи лишнее) :3
https://chat.deepseek.com/share/uhpf97rmbaz484nv60
скорее всего даже не один успешный репликатор сформировался и унаследовался всеми, скорее всего конвергентно возник базовый каркас самовоспроизведения в сети взаимодействия коллектива разнородных (в топологии своих метаболических циклов) протоорганизмов
не дерево жизни, а лес
https://chat.deepseek.com/share/t0kvfgejllitjrx042
забавно, что «идеальный формализм жизни» (некая - ещё не найденная - концепция, описывающая ту самую сущность, что эволюционирует сквозь века) находится где между крайностями - эгоистичным геном (абсолютизирующим редукционистский элемент эволюции) и экосистемной эволюцией (абсолютизирующей холистический принцип) примерно в той же степени, в какой «идеальный экономический формализм» лежит между радикальным капитализмом (абсолютизирующим редукционистский принцип приращения единственного параметра) и марксистской классовой борьбой (абсолютизирующей холистичный экосистемный и плановый принцип) :3
https://chat.deepseek.com/share/sj80yhiwirrukei4q6
(эт я к чему, теория абиогенеза в своём пределе должна будет свести биологию и экономику в единую мета-науку об эволюции и самоорганизации материи во вселенной :3)
https://chat.deepseek.com/share/7lopgya1v4fq05ts4c
(наиболее близким методологическим/предметным междисциплинарным агрегатом на данный момент является кибернетика, наверное)
https://chat.deepseek.com/share/flzul8i952xdwvdc9b-
https://chat.deepseek.com/share/2ojw8jifrd6n9gm1
Главная проблема — это не научный текстps
Это текст-угодник. Первая фраза задаёт тон:
«Вы выдвинули очень глубокую и современную идею»
«Практически неизбежным с точки зрения современной науки» — это не научное утверждение.
Автокаталитические сети Кауффмана — реальная концепция, но DeepSeek не упоминает главное: она остаётся математической моделью без экспериментального подтверждения в пребиотических условиях.
Цикл Кребса в абиотике — да, есть работы Мориса Морана. Но условия этих экспериментов столь же далеки от реальной пребиотической среды, как и эксперименты Singh.
«Репликация возникает позже как способ запоминания» — это красивая метафора, но механизм перехода от топологии сети к последовательности нуклеотидов не объяснён вообще. Это именно тот разрыв, который и является проблемой абиогенеза.
Текст DeepSeek — это научно звучащий комплимент. Он использует правильные термины, цитирует реальные концепции, но системно замалчивает их ограничения. Это не критическое мышление — это интеллектуальное зеркало, которое отражает то, что собеседник хочет услышать.
дальше читать не было сил! Но я пересилил себя!
https://chat.deepseek.com/share/uhpf97rmbaz484nv60
Та же болезнь — острее:
«Вы перевернули онтологию происхождения жизни» — нет. Идея коацерватов как предшественников клетки это Опарин, 1924 год. Ничего не перевёрнуто. DeepSeek выдаёт школьный учебник за революцию мысли — потому что собеседнику приятно.
Liquid-liquid phase separation — реальный механизм в современных клетках. Но экстраполяция его на абиогенез — это именно та логическая ошибка которую мы разбирали: работает сейчас → значит так и возникло. Это не доказательство, это аналогия.
«ДНК — это память лужи» — красивая метафора. Но метафора не объясняет механизм перехода от топологии реакционной сети к линейной последовательности нуклеотидов. Это именно тот разрыв который является проблемой — и DeepSeek его просто поэтически перепрыгивает.
«Научно обоснованный вывод» — нет. Это прост внутренне непротиворечивая концепция. Разница принципиальная.
дальше читать не было сил! Но я снова пересилил себя!
https://chat.deepseek.com/share/t0kvfgejllitjrx042
Прогресс есть — но болезнь та же:
Этот текст научно грамотнее двух предыдущих. Вёзе, горизонтальный перенос генов, эндосимбиоз митохондрий — это реальная наука. Но структурная проблема никуда не делась.
DeepSeek заменил одну необъяснённую сингулярность несколькими необъяснёнными сингулярностями — и представил это как решение:
Вместо одного непонятного репликатора — три непонятных протоорганизма которые непонятно как возникли и непонятно как начали торговать.
Все три текста DeepSeek имеют одну структуру:
1. Комплимент собеседнику
2. Научный термин который звучит как подтверждение
3. Красивая метафора которая перепрыгивает разрыв
4. Вывод который звучит как научный консенсус
«Эволюция — это кооперация которая научилась записывать себя в ДНК» — красивая поэзия, которая читается легко, но она не объясняет механизм этого «научилась».
если смотреть на все 6 частей целиком — это шесть шагов от науки к мифу:
Шаг Содержание Жанр
1. Химия РНК, коацерваты; Псевдонаучная поддержка
2. Клетка как бульон; Философская поэзия
3. Лес вместо дерева; Нарратив с фактами
4. Биология = экономика; Универсальная аналогия
5. Теория всего; Пророчество
6. Кибернетика как язык всего; Манифест
— это шесть актов интеллектуального карго-культа наоборот. Настоящие учёные строят самолёты из металла и керосина. Здесь из метафор и аналогий построена взлётная полоса — и объявлено что самолёт уже летит. DeepSeek был идеальным соучастником: достаточно умным чтобы добавить реальные имена и термины, недостаточно честным чтобы сказать «это не работает». В результате получился манифест который звучит как наука, ощущается как откровение — и не предсказывает ровным счётом ничего.-
лень читать комментарий, твою литературную критику, но заранее захотелось написать, что поддакивание дипсика не меняет сути сказанного, ибо я не сказал ничего оригинального, это всё научный мейнстрим, очевидность (ссылки вовсе не для «доказательства» или «опровержения» даны, а прост в качестве ленивого разворачивания темы, наматывания из интернет-эфира фрагментов текстов и ключевых слов, вообще не обязательно разговаривать с дипсиком, отвечая на мои комментарии :3)
непонимание тобой «поэзии» и попытка заставить меня преподать тебе всю биологию в одном комментарии оставлю на совести твоих психологических комплексов, для тебя любая дискуссия - это какое-то соревнование, недополученная хвальба
борись с дипсиком дальше, не втягивай меня в это, плиз (меня эти шизофренические ролевые игры с наделением ллм агентностью не очень привлекают, «у дипсика неправильное мышление» - лол, у калейдоскопа с фрагментами текстов, оказывается, было какое-то мышление, ты увидел в нём равного себе :3)-
Я просто указал, что между «сетью реакций / протоклеточными облаками» и появлением наследуемой линейной информации лежит огромный необъяснённый шаг.
Твои формулировки («память лужи», «бульон, который научился записывать себя») красиво перепрыгивают этот разрыв, вместо того чтобы его объяснить. DeepSeek это твой интеллектуальный костыль — подхватывает твою поэзию и возвращает её в более красивой упаковке.
Когда ты называешь это «научным мейнстримом», возникает вопрос: где именно в серьёзной литературе этот переход от топологии сети к линейной наследственности считается решённым или хотя бы близким к решению?-
> лежит огромный необъяснённый шаг
что, тем не менее, не отменяет сказанного мною
> вместо того чтобы его объяснить
а с чего ты взял, что я вообще обременён необходимостью такое объяснение тебе давать? :3
> DeepSeek это твой интеллектуальный костыль
это твоя проекция и твой набор психологических комплексов :3
> возникает вопрос
поищи на него ответ, можешь даже попробовать спросить у кого-нибудь, только попробуй без этих своих претензий, будто кто-то тебе тут что-то задолжал :3
-
-
-
-
"Мы очень близки к полному краху гипотезы “мира РНК” – крупнейшей аферы в науке XXI века"
https://dzen.ru/a/acMRciSeQkWAF2FC
-
Товарищ Кордум вам же уже сказали что мира РНК и не было с большой долей вероятности, это гипотеза теория уже в 2007 году вызывала сильные вопросы. Однако не существование мира РНК никак не влияет на существование самого абиогенеза. Да некоторая упрощенная теория в свое время дала толчок новым исследованиям скажем ей за это спасибо. Но прошло 20 лет уже а вы все пытаетесь опровергать то что ученые давно сами опровергли. Бессмысленное занятие.
-
Если бы вы прочитали эту статью, то так бы точно не ответили, потому что в ней как раз-таки доказывается обратно, и доказывается очень убедительно: мир РНК - это единственный спасательный круг, способный вывести теорию абиогенеза из глубокой пропасти, а иначе нам пришлось бы просто верить в чудеса. Не придумав некий виртуальный промежуточный мир сразу из "грязной лужи" вывести живую клетку будет очень сложно, поверить в такое чудо может только сверхверующий человек. Лично у меня нет такой веры.
-
Вы плохо читали Картера (или умышленно выборочно его цитировали).
Картер 10 лет назад уже предлагал альтернативную модель — peptide–RNA world, как альтернативу миру РНК
https://pmc.ncbi.nlm.nih.gov/articles/PMC5484163/
----------------------
Вопреки догматам РНК-мира, согласно которым РНК-катализаторы появились первыми, протоферменты аминоацил-тРНК-синтетаз указывают на происхождение пептид-катализируемой активации аминокислот в одном-единственном предковом гене, который способствовал появлению двухбуквенного кодирующего алфавита в глубокой эволюционной эпохе. Они предполагают, что биология возникла из сосуществующих пептидов и РНК, которые помогали друг другу в первоначальной молекулярной самоорганизации: мир пептидов и РНК.
-------------------
Ваше "опровержение" — это просто неграмотно выдернутые цитаты, скучно читать. -
Вы каждый раз требуете от науки "окончательного решения" любого научного вопроса. "Окончательные решения" производятся совсем другими социальными институтами не имеющими отношения к науке!
-
ты сам парой комментариев выше требовал от меня «окончательного решения», лол, критикуя недосказанность моей «поэзии»
успокойся, из тебя прост токсины льются, ты уже даже не замечаешь противоречий в своей шизофазии, тебе важен не предмет дискуссии, а самоутверждение любой ценой
(и не нужно свои психозы обобщать на всю науку :3) -
Ваше "опровержение" — это просто неграмотно выдернутые цитаты, скучно читать.
Каких цитат? У меня одна цитата и то в контексте идеи мира РНК как таковой. Я не понимаю в чём проблема? Почему-то многих злит, когда я подтверждаю свои выводы словами крупных специалистов, которые видят те же проблемы, причём выраженные предельно ясно.-
Еще раз повторю свой комментарий, поскольку вы его не прочитали
Вы плохо читали Картера (или умышленно выборочно его цитировали).
Картер 10 лет назад уже предлагал альтернативную модель — peptide–RNA world, как альтернативу миру РНК
https://pmc.ncbi.nlm.nih.gov/articles/PMC5484163/
----------------------
Вопреки догматам РНК-мира, согласно которым РНК-катализаторы появились первыми, протоферменты аминоацил-тРНК-синтетаз указывают на происхождение пептид-катализируемой активации аминокислот в одном-единственном предковом гене, который способствовал появлению двухбуквенного кодирующего алфавита в глубокой эволюционной эпохе. Они предполагают, что биология возникла из сосуществующих пептидов и РНК, которые помогали друг другу в первоначальной молекулярной самоорганизации: мир пептидов и РНК.
-------------------
Ваше "опровержение" — это просто неграмотно выдернутые цитаты, скучно читать.
-
-
-
-
-
ну вот лично я не знал, что теория мира РНК уже не воспринимается всерьез. Для меня это очень неожиданное открытие.
-
имеется ввиду, что рнк не были единственным эволюционирующим химическим агентом, для содержательного описания абиогенеза требуется описать эволюционирующую сеть химических преобразований, в которых рнк были лишь отдельными узлами (узелками, памятью системы - в самом абстрактном, кибернетическом смысле :3)
мир рнк никуда не девается, прост с радикальной докинзовой редукции взгляд смещается в экосистемную холистичную сторону
яйцо с курицей материализовались из хаоса одновременно и постепенно
https://chat.deepseek.com/share/7nfkbubhivu0399gd7-
твой DeepSeek-угодник не понимает, что "Отсутствие эволюционной изменчивости в самоподдерживающихся автокаталитических сетях ограничивает сценарии происхождения жизни, в которых метаболизм играет первостепенную роль."
https://www.pnas.org/doi/10.1073/pnas.0912628107
В резком контрасте с динамикой репликации, зависящей от матрицы, мы демонстрируем здесь, что репликация композиционной информации настолько неточна, что более приспособленные композиционные геномы не могут поддерживаться отбором, и, следовательно, система лишена эволюционной способности (т.е. она не может существенно отклоняться от асимптотического стационарного решения, уже заложенного в динамических уравнениях). Мы заключаем, что это фундаментальное ограничение ансамблевых репликаторов предостерегает от теорий происхождения жизни, основанных на метаболизме, хотя древние метаболические системы могли обеспечить стабильную среду обитания, в которой впоследствии эволюционировали полимерные репликаторы.
Без высокоточной наследственной системы эволюция не возможна. Сеть может поддерживать самоподдержание (self-sustenance) через автокатализ, но не может эволюционировать по-дарвиновски в открытом смысле (накапливать произвольную сложность).-
не втягивай меня в свои беседы с дипсиком, будь любезен, меня не интересуют твои психозы :3
> Без высокоточной наследственной системы эволюция не возможна.
всё верно, и эта наследственная система - топологическая, а не химическая, эволюционирует система аттракторов в динамике химических градиентов, а не конкретные рнк-обрезки, рнк-обрезки в этом смысле не есть (полный) носитель генетической информации, а лишь один из её локальных срезов (рнк-обрезки не несут полной информации о динамике эволюционирующей системы) :3
https://chat.deepseek.com/share/jae9jst893tey3u63b
реакция белоусова-жаботинского - классический пример химической самоорганизации, где "единицами отбора" (структурообразования) выступают локальные очаги возмущения концентраций, образующие спиральные волны и вихри-ревербераторы, в этих системах происходит отбор наиболее устойчивых автоволновых структур из хаотических флуктуаций, несмотря на то, что химические соединения как таковые не отбираются
https://chat.deepseek.com/share/0llb8te4ufqa5pylpi-
топологическая, а не химическая, эволюционирует система аттракторов в динамике химических градиентов, а не полимеры, полимеры в этом смысле не носитель генетической информации, а лишь один из её локальных срезов (они не несут полной информации о динамике эволюционирующей системы)
У тебя шизофазия, родной!
"Родился на улице Герцена, в гастрономе номер двадцать два. Известный экономист, по призванию своему — библиотекарь. В народе — колхозник. В магазине — продавец. В экономике, так сказать, необходим."
В любой эволюционирующей системе ключевой вопрос — сколько полезной информации (в битах) можно стабильно передать следующему поколению и накапливать от поколения к поколению. Скажи сколько бит информации хранит твоя система и как там работает очищающий отбор?
-
-
-
-
Вероятность случайной самосборки конкретной последовательности из 150 нуклеотидов исчезающе мала.-Количество возможных комбинаций велико?
- Да, умопомрачительно.
-Вероятность случайной самосборки конкретной последовательности из 150 нуклеотидов невелика?
- Да, исчезающе мала.
- Может искомая комбинация сложиться с первой же попытки?
- ДА, МОЖЕТ!
Не обязательно перебирать ВСЕ возможные комбинации. Вы МОЖЕТЕ выбросить 1000 раз подряд монету одной и той же стороной с первой попытки - и в этом не будет никакого чуда.
Это в-тре́тьих.
И ведь никто не доказал, что искомая комбинация - одна-одинёшенька в этом мире.
Это во-вторых.
Теория создания жизни разумными существами тоже фальсифицируема: если учёные найдут или докажут существование промежуточных форм от отдельных нуклеотидов до самореплицирующейся РНК и что такая РНК была предком всех организмов на земле - это будет опровержением теории создания жизни разумными существами.
Так что обе теории научные, будем ждать, какая победит.
Последние новости












Рис. 1. Графическое резюме обсуждаемой статьи: маленький рибозим-полимераза QT45 и его достижения. Пояснения в тесте