Инфракрасное излучение — древнейший сигнал для привлечения опылителей

Обычно мы представляем энтомофильное опыление как игру красок и ароматов: яркие цветки и сладкие запахи привлекают насекомых. Однако некоторые растения используют куда менее очевидный сигнал — тепло. В недавней статье в журнале Science показано, что инфракрасное излучение, возникающее при нагреве репродуктивных органов растений, служит полноценным сигналом для опылителей. Тепло, испускаемое в виде инфракрасного излучения, привлекает жуков, которые ориентируются на него с помощью специализированных сенсоров на усиках.
Растения за миллионы лет изобрели поразительно богатый арсенал способов привлечь опылителей. Цвет, запах и даже влажность давно считаются классическими сигналами для привлечения насекомых и других животных. Но некоторые растения, чтобы приманить опылителей, нагреваются. Как и у теплокровных животных, тепло у таких растений возникает как побочный продукт клеточного дыхания. Самые усердные из них работают с такой интенсивностью, что по уровню метаболизма могут сравниться с колибри в полете. В результате температура тканей растения может быть более чем на 35°C выше температуры окружающей среды.
Растительный термогенез (см. Термогенные растения) связывают с защитой от заморозков, усилением испарения ароматических веществ и даже созданием теплого убежища для опылителей. Однако есть основания полагать, что тепло само по себе может служить сигналом. Дело в том, что термогенез у растений возникает исключительно в репродуктивных органах — шишках у голосеменных и цветках у покрытосеменных, причем температурные паттерны там столь же разнообразны, как окраска лепестков.
Тепло от цветков и шишек может распространяться разными способами: через теплопроводность, конвекцию или инфракрасное излучение. В отличие от первых двух способов, инфракрасное излучение не требует прикосновения и распространяется во все стороны, словно невидимое сияние. Для животных это вовсе не экзотика: многие из них отлично «видят» тепло. Змеи, летучие мыши-вампиры, кровососущие комары и постельные клопы используют инфракрасное излучение, чтобы находить своих теплых жертв, а жуки-пирофилы с его помощью находят лесные пожары.
Учитывая, что большинство термогенных растений начинают нагреваться в сумерках и опыляются ночью, когда инфракрасное излучение особенно заметно, возникает логичный вопрос: а не используют ли опылители тепло как сигнал к «накрытому столу»? Именно это и стало предметом исследования статьи, недавно опубликованной в Science (см. W. A. Valencia-Montoya et al., 2025. Infrared radiation is an ancient pollination signal).
Авторы сосредоточились на опылении саговниковых — группы семенных растений, более близкой к хвойным, чем к покрытосеменным, и значительно более древней (см. «Живые ископаемые» саговники оказались вовсе не такими старыми, «Элементы», 16.12.2011). Современные саговниковые в основном встречаются в тропиках и образуют деревянистые стволы, крупные кожистые листья и репродуктивные структуры, напоминающие большие сосновые шишки. Саговники — древнейшая из ныне живущих линий семенных растений, опыляемых животными. На их долю приходится более половины всех известных термогенных видов растений. Мужские и женские репродуктивные органы формируются на разных растениях и имеют разные суточные циклы выделения летучих веществ, заставляя жуков-опылителей курсировать между мужскими (пыльцевыми) и женскими (семенными) шишками (см. Австралийские саговники управляют насекомыми-опылителями при помощи кнута и пряника, «Элементы», 10.10.2007).
Палеонтологическая летопись подсказывает, что эти отношения — не мимолетный роман, а давний союз: судя по окаменелостям, взаимодействие саговников и жуков-опылителей существует как минимум с ранней юры, около 200 миллионов лет назад. Именно поэтому саговники и жуки стали идеальной модельной системой, помогающей разобраться в роли теплового инфракрасного излучения в коммуникации между растениями и их опылителями — и заглянуть в самые истоки эволюции опыления.
Сначала исследователи обратились к тепловизору. С его помощью они получили термические изображения саговника замия шелушащаяся (Zamia furfuracea) и подтвердили наличие «нагревательного эффекта» в пылящих мужских шишках (рис. 1). Именно эти структуры, а не всё растение целиком, работают как локальные тепловые генераторы.
Термогенез у растений жестко завязан на окислительное фосфорилирование — метаболический путь, при котором энергия, образовавшаяся при окислении питательных веществ, запасается в митохондриях клеток в виде АТФ. Однако при термогенезе из-за действия некоторых ферментов, таких как альтернативная оксидаза (AOX), углеводы или липиды обходят стандартный энергетический путь и идут не на синтез АТФ, а буквально «сжигаются впустую», рассеивая окислительно-восстановительную энергию в виде тепла. Иными словами, митохондрии временно переключаются с роли электростанции на роль обогревателя.
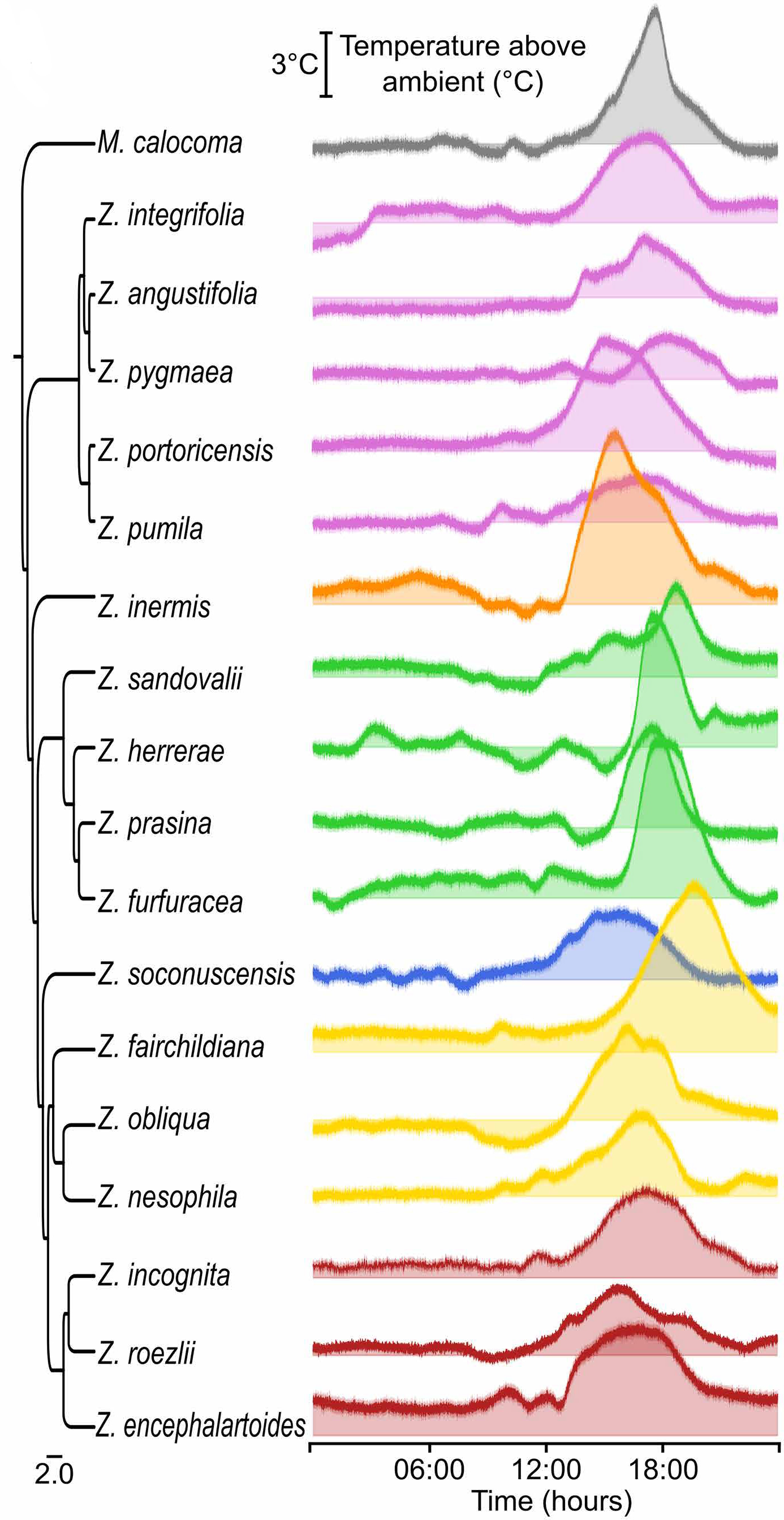
Рис. 2. Превышение температуры шишек над температурой окружающей среды в зависимости от времени суток (по оси х отложено время в часах) для 17 видов рода Zamia, охватывающих все основные эволюционные линии этого рода (всего описано 89 видов), а также для близкородственного вида Microcycas calocoma, использованного как внешняя группа для построения дерева. Рисунок из обсуждаемой статьи
В шишках Z. furfuracea исследователи обнаружили заметно больше митохондрий, чем в нетермогенных тканях. Более того, непосредственно перед началом нагрева и во время него резко возрастала активность гена AOX1. Параллельно усиливалась экспрессия генов, связанных с транспортом углеводов и метаболизмом крахмала. В клетках спорофиллов наблюдалось множество амилопластов, плотно набитых крахмалом, которые после фазы термогенеза практически опустошались. Всё это указывает на то, что именно AOX1 играет ключевую роль в управлении дыханием у саговников, а главным «топливом» для выработки тепла служит крахмал.
Когда исследователи проследили динамику нагрева во времени, стало ясно: термогенез подчиняется строгому биологическому расписанию. Каждый день происходил один мощный «тепловой всплеск», начинавшийся после полудня и достигавший пика ранним вечером. Чтобы понять, насколько это явление универсально, авторы расширили анализ и изучили термогенез у разных представителей рода Zamia — самой разнообразной неотропической группы саговников. Результат оказался однозначным: все исследованные виды оказались термогенными, и у всех выработка тепла подчиняется расписанию (рис. 2).
Таким образом, исследователи пришли к выводу, что термогенез у Zamia — это эволюционно сохраненный, энергозатратный и жестко синхронизированный по времени процесс, который проявляется исключительно в органах, участвующих в опылении. Проще говоря, растение греется не «вообще», а строго по расписанию и строго там, где это действительно нужно.
Саговники — растения раздельнополые: мужские и женские шишки развиваются на разных особях. Исследователи обнаружили, что у всех изученных видов Zamia «мальчики» и «девочки» живут по разному тепловому расписанию. Суточная динамика нагрева у мужских и женских растений заметно отличалась. У Zamia furfuracea мужские шишки начинали нагреваться раньше: они быстро выходили на пик температуры и затем остывали. Женские же шишки включались в процесс примерно на три часа позже, достигая максимального нагрева уже после того, как мужские успевали «остыть».
Такое рассогласование выглядело слишком точным, чтобы быть случайным, и побудило авторов исследовать взаимосвязь между термогенезом растений и поведением опылителей. Они проследили за поведением жука Rhopalotria furfuracea — главного опылителя Z. furfuracea. В полевом эксперименте жуков пометили флуоресцентными красителями, светящимися в ультрафиолете, что позволило буквально «подсвечивать» маршруты насекомых (рис. 3). Оказалось, что жуки тянулись прежде всего к самым теплым участкам мужских шишек, показывая, что тепловые узоры шишек могут служить для опылителей своеобразными дорожными указателями.

Рис. 3. (A) Мужские шишки саговника Zamia furfuracea в момент выброса пыльцы. Пыльца была помечена ультрафиолетово-флуоресцентными красителями, чтобы отслеживать перемещения жуков-опылителей. (B) Жук-опылитель (Rhopalotria furfuracea), покрытый пыльцой и флуоресцентными метками, во время переноса пыльцы с мужских растений на женские. (C) Женская шишка Z. furfuracea с заметными следами флуоресцентного красителя, оставленными жуками-опылителями в процессе переноса пыльцы. Рисунок из обсуждаемой статьи
Эксперименты в контролируемых условиях это подтвердили. В клетках, где можно было точно отслеживать перемещения насекомых, присутствие жуков на мужских и женских шишках четко совпадало с подъемами и спадами температуры, причем отдельно для каждого пола (рис. 4). Более того, исследователи выдвинули гипотезу, что именно охлаждение мужских шишек служит сигналом «пора уходить» — побуждая жуков покинуть пыльцевые шишки и перелететь к женским, готовым принимать пыльцу.
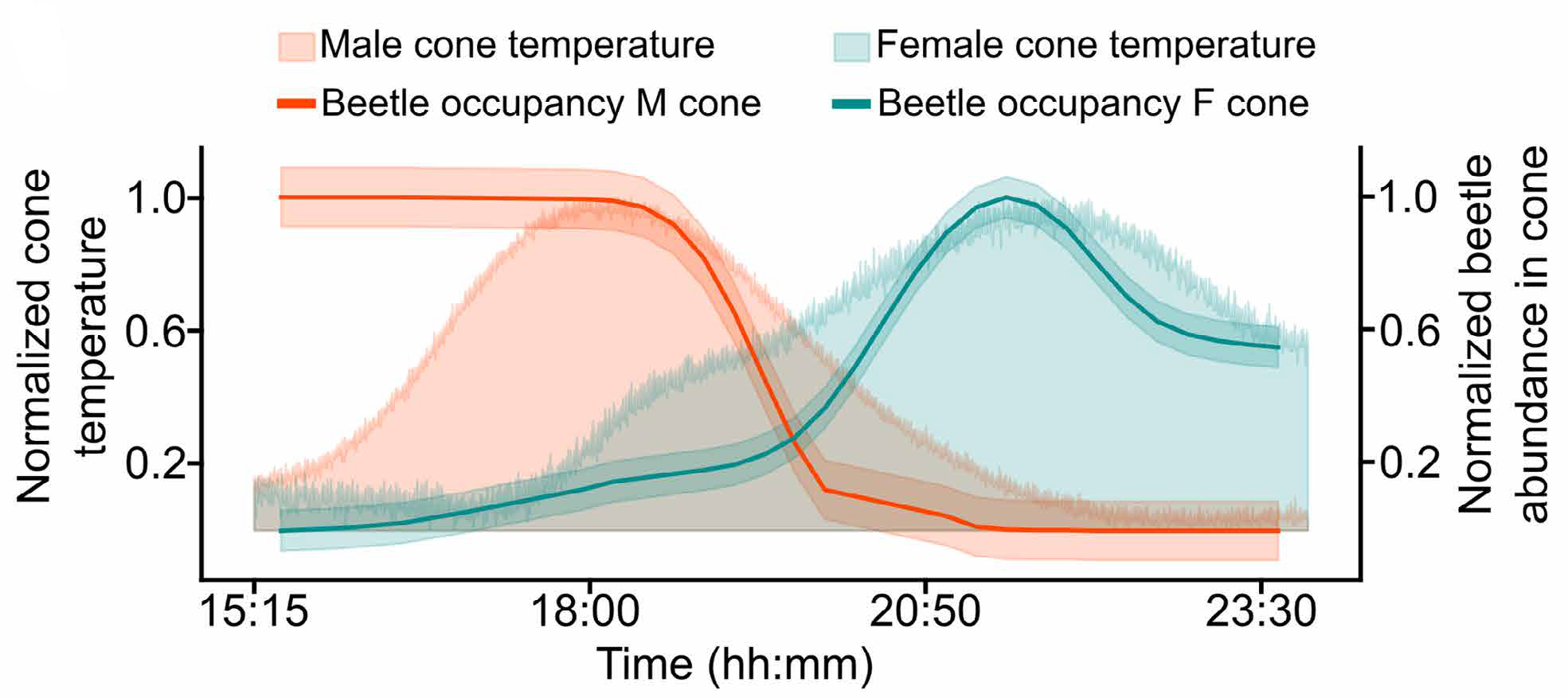
Рис. 4. Связь между нагревом шишек и поведением жуков-опылителей во времени. По горизонтальной оси показано время суток (часы:минуты). Левая вертикальная ось — нормализованная температура шишек, правая — нормализованная численность жуков на шишках. Оранжевая заливка — температура мужских шишек, зеленая — температура женских шишек. Оранжевая линия — присутствие жуков в мужских шишках, зеленая линия — присутствие жуков в женских шишках. Вечером мужские шишки активно нагреваются, и жуки концентрируются на них. По мере снижения температуры мужских шишек и одновременного нагрева женских жуки покидают мужские шишки и переходят в женские, обеспечивая перенос пыльцы. Рисунок из обсуждаемой статьи
Ранее считалось, что у саговников так называемый механизм «тяни-толкай» в опылении запускается главным образом летучими веществами, которые выделяют шишки. Однако запахи, как и влажность воздуха, часто меняются одновременно с нагревом. Чтобы отделить вклад тепла от всех остальных сигналов, авторы пошли на изящный экспериментальный трюк. Они отсканировали настоящие шишки и создали на 3D-принтере их трехмерные модели, которые можно было искусственно нагревать. Эти макеты размещали рядом с живыми растениями и нагревали либо до температуры, характерной для термогенеза, либо оставляли при температуре окружающей среды. Чтобы жуки не могли «пощупать» разницу, модели покрывали липким слоем без запаха: насекомое сразу прилипало при посадке и не использовало контакт для оценки температуры. Результат оказался однозначным. Жуки-опылители значительно чаще прилетали к нагретым моделям — как мужским, так и женским — чем к холодным (рис. 5). Это означает, что тепло само по себе работает как сигнал, действующий на небольших и средних дистанциях.

Рис. 5. (А) Полевые эксперименты с использованием напечатанных на 3D-принтере моделей женских шишек: слева — искусственно нагретая модель, справа — модель при температуре окружающей среды. (B) Посещения жуками-опылителями искусственных 3D-моделей шишек: нагретых (слева) и ненагретых (справа). (C) Доля жуков, посетивших нагретые (слева) и ненагретые (справа) модели мужских (M, снизу) и женских (F, сверху) шишек в полевых условиях. (D) Аналогичное сравнение в лабораторных экспериментах, где исключалось влияние теплопередачи через воздух и контакт (кондукция и конвекция). Рисунок из обсуждаемой статьи
Чтобы исключить влияние теплого воздуха и оставить только инфракрасное излучение, исследователи пошли еще дальше. Они накрыли нагретые модели полиэтиленовой пленкой, почти полностью прозрачной для инфракрасных лучей, но отсекающей конвекцию. И даже в этих условиях жуки предпочитали «инфракрасно светящиеся» шишки (рис. 5D). Это стало прямым доказательством того, что инфракрасное излучение — невидимое для нас, но вполне различимое для насекомых — действительно служит полноценным сигналом опыления. Иными словами, для жуков саговник — это не просто пахнущий цветок, а еще и теплый маяк, который буквально светит в инфракрасном диапазоне, направляя опылителей туда, где они нужнее всего.
Далее исследователи сделали логичный следующий шаг. Если саговники и их жуки-опылители действительно выработали в ходе коэволюции видоспецифичный канал связи, то у жуков должны быть специальные органы чувств, способные улавливать тепловое инфракрасное излучение. У насекомых за восприятие окружающего мира отвечают сенсиллы — микроскопические сенсорные структуры на поверхности тела. Их форма и расположение обычно напрямую связаны с функцией. Известно, что большинство температурочувствительных нейронов у насекомых находятся в так называемых целоконических сенсиллах типа «штырёк в ямке» — это крошечные углубления в хитине, внутри которых прячутся чувствительные отростки нервных клеток.
Авторы изучили антенны двух видов жуков — Rhopalotria furfuracea и Pharaxonotha floridana. Оба они опыляют термогенные саговники и потому были идеальными кандидатами для поиска «инфракрасного зрения». И действительно, у обоих видов самый кончик усика оказался буквально усыпан сенсиллами, форма которых наводила на мысль о термочувствительной функции. Чтобы убедиться, что это не просто совпадение, исследователи перешли от анатомии к физиологии. С помощью метода регистрации активности отдельных сенсилл они показали, что структуры на кончиках антенн P. floridana действительно реагируют на тепло. Причем реагируют по-разному. В одном типе целоконических сенсилл находился нейрон, который мгновенно «стрелял» в ответ на повышение температуры. В другом — сразу две группы нейронов: одни активировались при нагреве, другие, наоборот, подавлялись. Такая комбинация позволяет не просто чувствовать тепло, а тонко отслеживать его изменения. Для сравнения, волосковидные механорецепторы исправно реагировали на прикосновения, но оставались совершенно равнодушными к температуре.
Далее авторы измерили поведенческие реакции обездвиженных жуков на тепловое ИК-излучение. В качестве контрольного сигнала использовали знакомый и надежный ориентир — запах метилсалицилата, характерный для шишек. Жуки реагировали и на запах, и на инфракрасное излучение. Но после микроскопического удаления чувствительного кончика антенны происходило показательное разделение ролей: реакция на запах сохранялась, а вот отклик на ИК-излучение полностью исчезал (рис. 6).
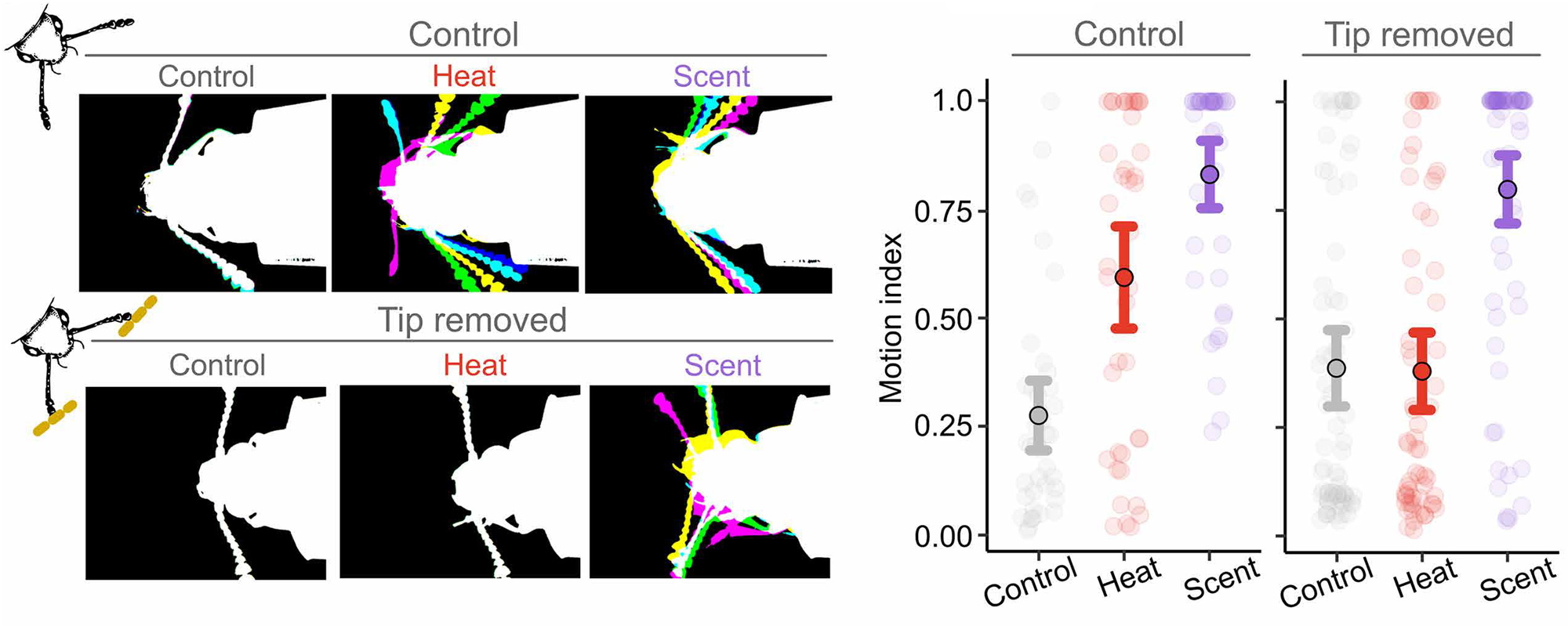
Рис. 6. Движения антенн жуков-опылителей в ответ на тепловое излучение (Heat) и запах шишек (Scent). Сравниваются контрольные жуки (Control) и жуки, у которых был удален последний сегмент антенны (Tip removed). Цвета антенн отражают движения в разные временные интервалы. На графике справа показано среднее значение и 95-процентный доверительный интервал индекса движения (ось y) в разных вариантах эксперимента. Рисунок из обсуждаемой статьи
Чтобы добраться до молекулярных основ инфракрасного «чутья» жуков, исследователи проанализировали транскриптом (то есть набор активно работающих генов) антенн жука Pharaxonotha floridana и сравнили его с другими органами. Картина оказалась показательной: именно в усиках резко повышалась активность генов, связанных с температурной чувствительностью. Среди них — GR28b (вкусовой рецептор, реагирующий на нагрев), IR21a (ионный рецептор, чувствительный к охлаждению) и несколько вариантов гена TRPA1 — хорошо известного «термодатчика» из семейства ионных каналов.
Дальнейшие эксперименты показали, что один из этих вариантов, TRPA1, работает строго в определенных сенсорных нейронах на самом кончике антенны — ровно в той зоне, которая необходима жуку для восприятия инфракрасного излучения. У некоторых насекомых, например у комаров, в восприятии инфракрасного излучения участвуют опсины — белки, обычно связанные со зрением. Однако у жуков авторы не обнаружили таких опсинов в антеннах.
TRPA1 давно известен как участник инфракрасного восприятия у змей и комаров, так что его появление в этой истории выглядело многообещающе. Особенно исследователей заинтересовал короткий вариант этого белка — TRPA1(B), который оказался самым активно экспрессируемым в антеннах жука. Чтобы проверить, действительно ли он работает как терморецептор, авторы экспрессировали этот белок в клетках и с помощью электрофизиологии измерили активность этого ионного канала — TRPA1(B) активно реагировал даже на очень слабое повышение температуры.
Затем последовал изящный фармакологический трюк. Ученые протестировали три десятка веществ, которые могли бы блокировать работу TRPA1. Большинство известных ингибиторов, эффективно работающих у позвоночных, мух и комаров, на жука не подействовали. Зато одно соединение — AM-0902 — оказалось точным «выключателем» жучиного TRPA1. Когда его наносили на антенны, жуки переставали реагировать на инфракрасное излучение, но по-прежнему чувствовали запахи шишек.
Но почему именно этот вариант TRPA1 так хорошо чувствует тепло? Чтобы разобраться, исследователи сравнили его с другим вариантом — TRPA1(A). Оказалось, что белки отличаются совсем немного: у TRPA1(B) просто укорочен N-конец молекулы. И всё же этого хватило, чтобы чувствительность к температуре выросла в разы. Тот же эффект обнаружился и у другого вида жуков-опылителей, Rhopalotria furfuracea, далекого родственника P. floridana.
Такие различия между вариантами TRPA1 известны и у мух, и у комаров, а новые данные показывают, что игра с вариантами этого гена началась еще как минимум 300 миллионов лет назад. Эволюционный анализ подтвердил: N-концевая часть TRPA1 — настоящий «горячий участок» адаптивных изменений, особенно у жуков, связанных с термогенными растениями.
Если TRPA1(B) действительно настроен на сигналы конкретного растения, то диапазон его активации должен совпадать с температурой шишек «своего» саговника. Именно это и обнаружилось. У P. floridana TRPA1(B) максимально активно реагировал на температуры, характерные для его хозяина — Zamia integrifolia. У R. furfuracea диапазон был другим и точно соответствовал температурным пикам Zamia furfuracea (рис. 7). Эти жуки-опылители относятся к семействам, разошедшимся более 140 миллионов лет назад, но оба ориентируются по ИК-сигналам. Это говорит о том, что восприятие ИК-сигналов — вовсе не экзотика для охотников и паразитов, как считалось раньше, а вполне рабочий и эволюционно успешный канал общения между растениями и их опылителями.

Рис. 7. (A) Профили термогенеза у растений, опыляемых разными видами жуков: Zamia integrifolia (опыляется P. floridana, более низкие температуры, розовый цвет), Zamia furfuracea (опыляется R. furfuracea, более высокие температуры, зеленый цвет). (B) Температурные ответы ионных каналов TRPA1(B), выделенных у жуков P. floridana (розовый) и R. furfuracea (зеленый). Рисунок из обсуждаемой статьи
Цветки и шишки растений можно сравнить с рекламными щитами: они используют сразу несколько сигналов, чтобы привлечь опылителей. Эти сигналы работают на разных расстояниях и в разное время. Так, запах — один из самых древних и универсальных способов привлечения — может заманивать насекомых с расстояния в несколько метров. Однако на близкой дистанции одного запаха недостаточно: чтобы точно направить опылителя к источнику пыльцы или нектара, нужны дополнительные, «короткодистанционные» сигналы, например окраска.
Исследователи обнаружили, что инфракрасное (тепловое) излучение эффективно именно на ближних и средних расстояниях и служит сигналом, указывающим на шишки, которые выделяют или принимают пыльцу. Поскольку в этих диапазонах у современных растений доминируют цветовые сигналы, авторы задались вопросом: дополняет ли цвет тепловые сигналы или является избыточным? Чтобы ответить на него, они сравнили распространение тепловых и цветовых сигналов у разных групп семенных растений.
Если самые ранние системы «растение — опылитель» действительно опирались не на цвет, а на метаболические сигналы вроде тепла, то окраска цветков и шишек должна была быть слабо различимой для зрения древних опылителей. Чтобы проверить это, исследователи изучили цветовое зрение жуков — одних из самых древних опылителей. Анализ генов светочувствительных белков (опсинов) показал, что у них есть только два типа зрительных рецепторов: чувствительные к ультрафиолетовому и к длинноволновому (зеленому) свету. Это означает, что их зрение дихроматично и беднее по сравнению с трихроматическим зрением, характерным для многих современных насекомых.
Затем авторы проверили, насколько заметными являются цветовые сигналы растений для разных опылителей — жуков, пчел и бабочек. Они измерили спектры отражения света у шишек 23 видов саговников и сопоставили их с огромной базой данных по окраске цветков цветковых растений. Эти данные были «переведены» в модели цветового зрения соответствующих опылителей. Окраска шишек практически не совпадала с чувствительностью зрительных рецепторов жуков, то есть для них такие объекты визуально малозаметны. Напротив, цветки цветковых растений демонстрировали высокий цветовой контраст в моделях зрения пчел и бабочек — более поздних и визуально ориентированных опылителей. Это указывает на эволюционный сдвиг: от древних метаболических сигналов, таких как тепло, к всё более сложным и разнообразным цветовым сигналам, характерным для современных цветковых растений.
Чтобы проверить, насколько это правило общее, исследователи сопоставили распространение термогенеза и разнообразие окраски у основных семейств семенных растений. Выяснилось, что способность к термогенезу независимо возникала у саговников и покрытосеменных растений, но особенно часто встречается у ранних, эволюционно древних линий. В то же время более молодые группы, такие как розиды и астериды, отличаются богатством окраски и значительными метаболическими затратами на синтез пигментов (рис. 8). Статистический анализ показал выраженную отрицательную связь между термогенезом и цветовым разнообразием: растения, использующие тепловые сигналы, как правило имеют бедную окраску, и наоборот. Это указывает на эволюционный компромисс, связанный с тем, что разные сигналы частично дублируют друг друга, требуют разных энергетических затрат и нацелены на разные сенсорные предпочтения опылителей.
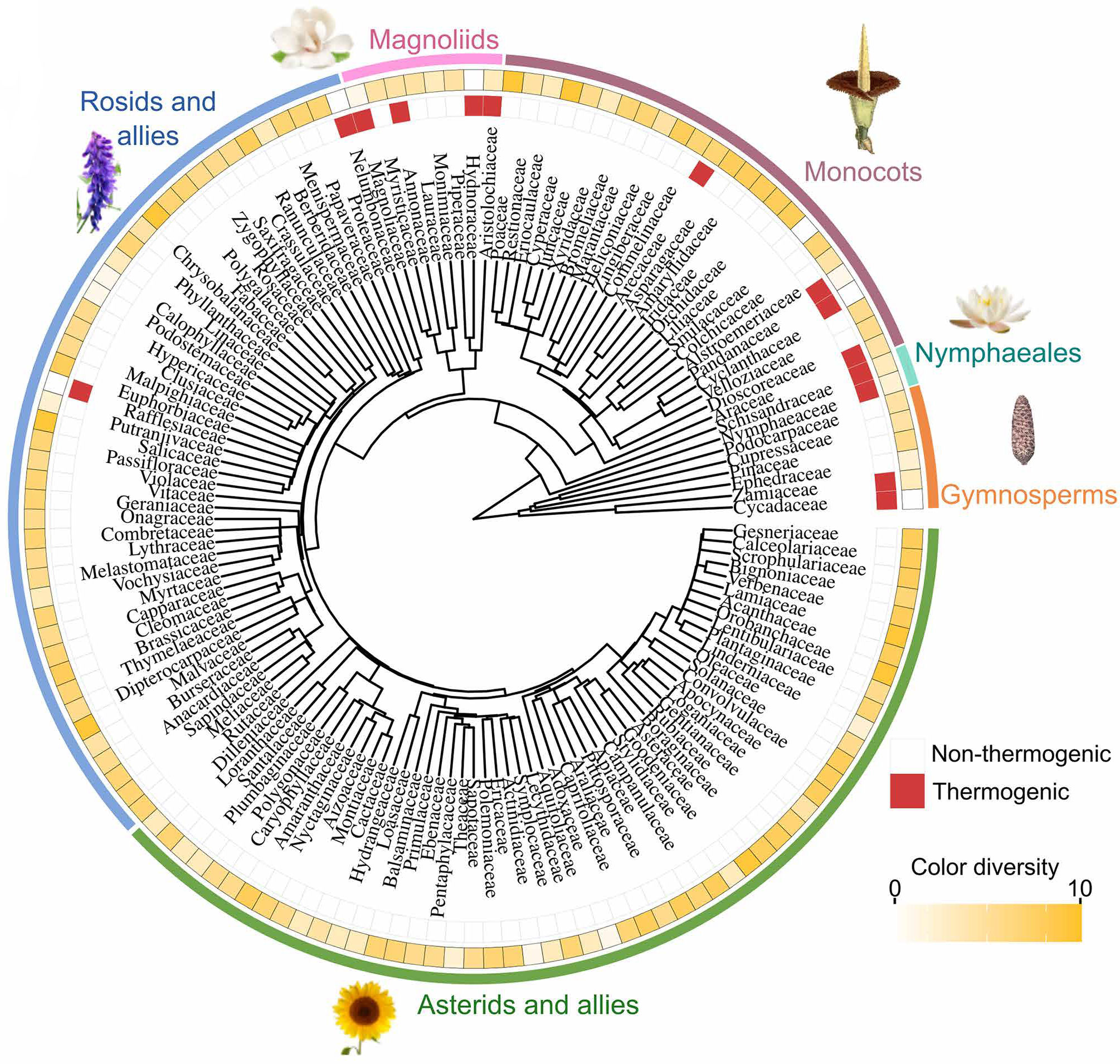
Рис. 8. Распределение термогенеза (красным отмечены термогенные растения, белым — нет) и разнообразия окраски (интенсивность желтого цвета отражает разнообразие окраски), нанесенное на филогенетическое древо семенных растений (Spermatophyta). Рисунок из обсуждаемой статьи
Наконец, авторы сопоставили данные о происхождении термогенеза с ископаемыми находками растений и насекомых. Анализ показал, что термогенез появился у саговников еще в раннем пермском периоде — около 275 миллионов лет назад — и представляет собой одну из самых древних известных стратегий опыления. Жуки, которые были ночными, обладали простым цветовым зрением и стали одними из первых опылителей семенных растений, возникли и начали диверсифицироваться раньше, чем пчелы и бабочки. Последние появились значительно позже и эволюционировали параллельно с бурным расцветом современных цветковых растений (рис. 9).
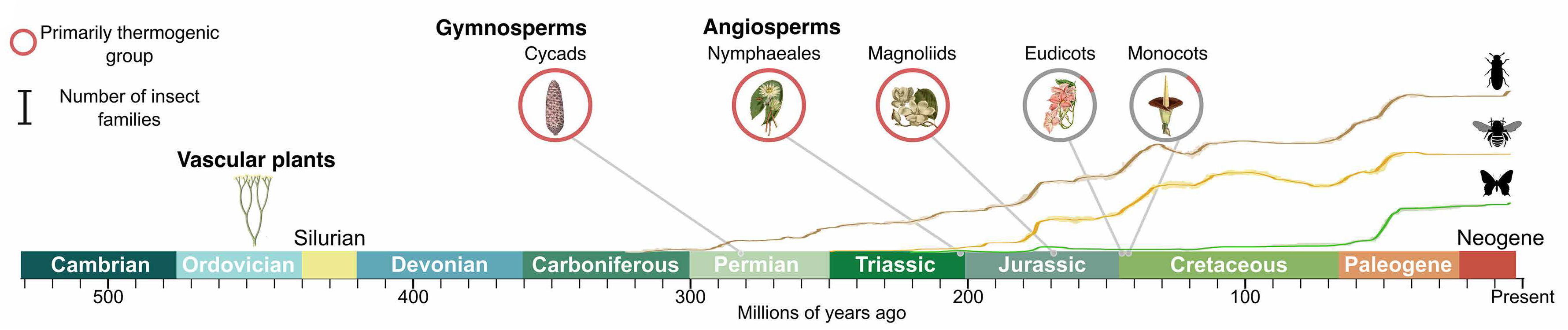
Рис. 9. Временная шкала эволюции основных групп растений и накопленного разнообразия семейств насекомых-опылителей в геологическом времени. По оси x миллионы лет назад. Красные круги означают группы, для которых характерен термогенез. Показаны три крупные группы насекомых: жесткокрылые (Coleoptera, жуки), перепончатокрылые (Hymenoptera: пчелы, осы, муравьи), чешуекрылые (Lepidoptera: бабочки и моли). Масштабная линейка соответствует 25 семействам насекомых. Рисунок из обсуждаемой статьи
Таким образом, ставка саговников на узкоспециализированный инфракрасный сигнал, эффективный, но бедный по «информационному содержанию», могла ограничить их эволюционный потенциал. В отличие от них, цветковые растения, освоившие практически безграничное пространство цветовых сигналов и дневных опылителей, получили ключ к своему исключительному разнообразию.
Источник: Wendy A. Valencia-Montoya, Marjorie A. Lienard, Neil Rosser, Michael Calonje, Shayla Salzman, Cheng-Chia Tsai, Nanfang Yu, John R. Carlson, Rodrigo Cogni, Naomi E. Pierce, and Nicholas W. Bellono. Infrared radiation is an ancient pollination signal // Science. 2025. V. 390. P 1164–1170. DOI:10.1126/science.adz1728.
Елена Устинова
-
-
Ну, вопросы не все закрыты, думаю... )
Полиэтилен меня только смутил.
Как с его помощью конвекции они думали избежать? Ведь, если инфракрасность излучается, то и воздух она греет.. Разве что трубу открытую сверху из него сделали столь высокую, чтоб поднимавшийся теплый воздух точно оказался не на пути полета опылителей-жуков...


Представил себя древним зверем и ощутил кайф такую шишку съесть: и тепленькая, и крахмала много - этакая допотопная шаурма!
да еще и бесплатно!
Жизнь и тогда уже была полна радости и удовольствий!
Вот вам еще один закон, еще одна основа жизни - делать другим приятно!)
А и то: кто бы за нее держался, если б удовольствий не было?))
-
Ну, если шкалу переписать так, что последний пункт будет нашими ушами иллюстрирован (правым ухом тише слыхать, значит надо налево"), то даже и не знаю... Ушами невероятно точно привык ориентироваться, при том как-то ими понимаю не только направление в горизонтальной плоскости, но и градус по отношению к горизонту безошибочно..
Плохого разрешения камера быть даже может и похуже ориентирует)
Последние новости





















Рис. 1. (A) Открытая шишка саговникового растения. (B) Тепловизионное изображение мужской шишки саговника в момент нагревания во время выброса пыльцы. Хорошо видно, что температура шишки заметно превышает температуру окружающей среды. Рисунок из обсуждаемой статьи