Гонка вооружений между деревом и долгоносиком запечатлена в их геномах

Дерево айлант высочайший и два его специализированных долгоносика ведут длящуюся много миллионов лет эволюционную «гонку вооружений», в которой каждая сторона совершенствует свой биохимический арсенал. Секвенирование геномов дерева и насекомых выявили у айланта дупликации генов, отвечающих за синтез защитных веществ, а у долгоносиков — расширение семейств генов, нейтрализующих эти яды и участвующих в восприятии сигналов окружающей среды. Их биоинформатический анализ показывает, как тесное трофическое взаимодействие может перестраивать геномы и направлять эволюцию обеих сторон.
Растения синтезируют сотни и тысячи вторичных метаболитов, способных отпугнуть или отравить насекомых-фитофагов. Эти соединения воздействуют на насекомых через самые разные биохимические механизмы: от нарушения работы нервной системы до подавления пищеварительных ферментов. Насекомые, в свою очередь, нередко развивают адаптации, позволяющие им нейтрализовать или обходить эти барьеры, и продолжают питаться на выбранных растениях-хозяевах, расти и размножаться. Взаимодействие растений и фитофагов рассматривается как динамичный коэволюционный процесс, часто описываемый метафорой «гонки вооружений» (см. например, Гонка вооружений — двигатель эволюции, «Элементы», 01.03.2010). Это эволюционное противодействие считается одним из ключевых факторов возникновения новшеств как в химическом арсенале растений, так и в физиологических и поведенческих адаптациях фитофагов, а также способствует взрывной диверсификации взаимодействующих видов.
В недавно опубликованной в журнале Molecular Ecology Resources статье исследователи из Китая и Австралии описали обнаруженные ими геномные особенности, связанные с возможной гонкой вооружений между айлантом высочайшим (Ailanthus altissima) и двумя специализированными долгоносиками, питающимися исключительно этим видом дерева.
Айлант высочайший (Ailanthus altissima) — быстрорастущее дерево, листья которого немного напоминают листья пальмы, а при растирании источают неприятный специфический запах. На своей родине, в северо-западном Китае, это дерево ценят за роль в создании полезащитных лесополос. Но, оказавшись на других континентах, айлант стал агрессивным захватчиком. Его успех объясняется не только быстрым ростом, но и высокой устойчивостью к насекомым-вредителям благодаря сочетанию широкого набора летучих веществ, вторичных метаболитов и уникальных для него квассиноидов (см. Quassinoid) и фенилпропаноидов. В результате дерево почти не повреждается фитофагами, и известно всего пять видов насекомых, способных питаться им. Два из них — «корневой» долгоносик Eucryptorrhynchus scrobiculatus и «стеблевой» долгоносик E. brandti. В Китае они представляют настоящую беду для айланта, а вот в Америке, где у айланта нет естественных врагов, их рассматривают как потенциальное оружие против инвазионного дерева. Эти долгоносики близки и по происхождению, и по образу жизни, причем питаются исключительно айлантом. Их можно встретить на одном и том же дереве, где они совместными усилиями наносят растению серьезный урон. Однако между ними есть четкое разделение труда (рис. 2): корневой долгоносик откладывает яйца в почву возле корней, а стеблевой — на ствол. Личинки первого повреждают корни, второго — ствол. Взрослые же особи обоих видов в симпатричных популяциях поедают стебли, побеги и черешки листьев.
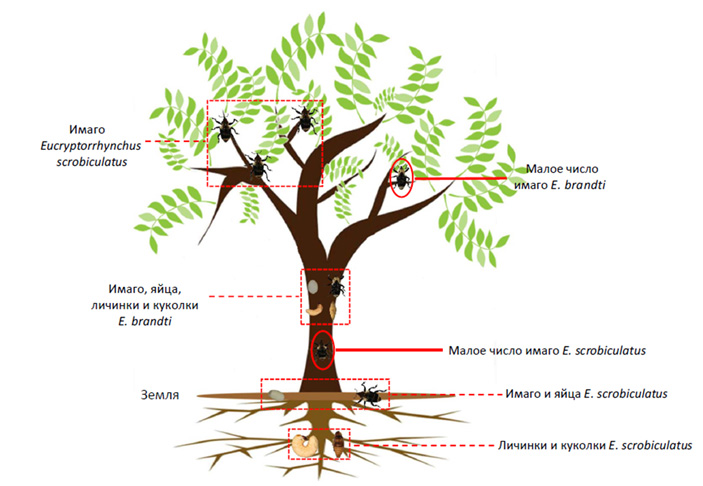
Рис. 2. Разделение ниш долгоносиков Eucryptorrhynchus scrobiculatus и E. brandti на айланте высочайшем. E. scrobiculatus предпочитают откладывать яйца на почву возле корней дерева-хозяина, а его личинки питаются корнями. E. brandti предпочитает откладывать яйца на ствол дерева, а его личинки питаются стволом. Рисунок из обсуждаемой статьи
Авторы обсуждаемой работы отсеквенировали геном айланта и обоих его специализированных долгоносиков, а затем сопоставили эти данные с транскриптомами и метаболомами. Целью было понять, какие генетические особенности обеспечивают успех каждой из сторон в этой химико-биологической дуэли. У айланта внимание сосредоточили на генах, отвечающих за синтез вторичных метаболитов. У долгоносиков — на генах рецепторов, воспринимающих сигналы окружающей среды, и ферментов, участвующих в детоксикации. Ученые не только описали состав сложных вторичных метаболитов айланта, но и аннотировали гены насекомых, потенциально отвечающие за нейтрализацию токсинов растения.
Генетическое сравнение показало, что айлант и его ближайший родственник Toona sinensis разошлись эволюционными путями около 29 миллионов лет назад. Долгоносики, специализирующиеся на айланте, разошлись между собой еще раньше — примерно 39 миллионов лет назад. История численности их популяций (имеется в виду эффективная численность) удивительным образом совпадала с историей айланта: у корневого долгоносика эффективная численность была стабильно ниже, тогда как у стеблевого колебания шли в том же ритме, что и у дерева, но с небольшим временным сдвигом (рис. 3). Это может говорить о давней экологической связи насекомых с предками айланта.
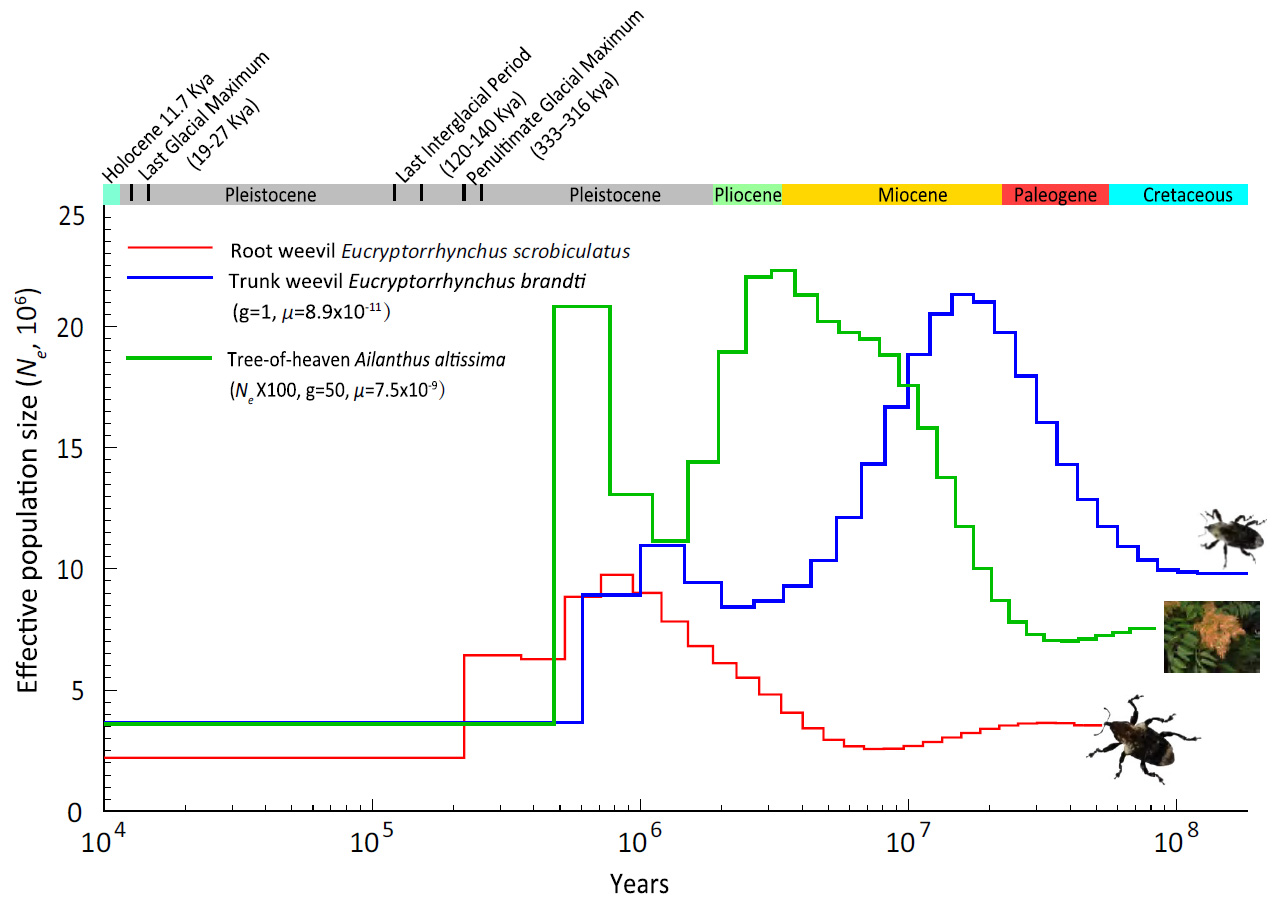
Рис. 3. Динамика эффективной численности популяций айланта (Ailanthus altissima, зеленая линия) и двух специализированных долгоносиков — Eucryptorrhynchus scrobiculatus (красная линия) и E. brandti (синяя линия) — за десятки миллионов лет. По горизонтальной оси — время в логарифмическом масштабе (наше время — слева), по вертикальной оси — эффективная численность (×10⁶). Цветные полосы вверху указывают геологические эпохи, а над ними отмечены ключевые климатические события позднего плейстоцена и голоцена. Видно, что у E. brandti колебания эффективной численности происходили в том же ритме, что и у айланта, но с временным сдвигом, а у E. scrobiculatus эффективная численность была более низкой. Рисунок из обсуждаемой статьи
Когда исследователи сравнили хромосомы айланта между собой и с геномом T. sinensis, они заметили, что каждый крупный участок ДНК представлен в двойном экземпляре. Такой «дубль» — признак древнего события полного удвоения генома. По скорости накопления молекулярных изменений ученые рассчитали, что произошло это относительно недавно, около 14 миллионов лет назад, уже после расхождения с T. sinensis. Удвоение генома дает растению дополнительный набор генов, часть из которых может со временем изменяться и приобретать новые функции, в том числе — связанные с защитой (см. Многофункциональные гены — основа для эволюционных новшеств, «Элементы», 30.06.2008).
Геном айланта оказался обширным — было выявлено 43 139 белок-кодирующих генов. Почти 2800 из них заметно «разрослись» по сравнению с другими деревьями порядка Sapindales. Многие из этих расширенных семейств участвуют в синтезе вторичных метаболитов — химических соединений, которые не нужны растению непосредственно для роста, но могут использоваться для защиты от фитофагов. Особенно выделяются гены, связанные с производством квассиноидов — редких для растений веществ с выраженным токсическим и антипищевым действием. Часто эти гены встречаются группами дубликатов, что, вероятно, связано с давним удвоением генома. Авторы предполагают, что именно такие дупликации ключевых генов и определяют химическое разнообразие квассиноидов, а значит — и эффективность защиты айланта от насекомых.
Химический анализ подтвердил: в тканях айланта нашли 745 соединений из 11 классов, и больше половины из них — вторичные метаболиты. Квассиноиды оказались чемпионами по разнообразию. Лидером по количеству стал айлантон: в корнях его почти в десять раз больше, чем в стволе. Амаролида в корнях тоже много, тогда как айлантинол и синджугликозид концентрируются в стволах. Такое распределение соединений между корнями и стволом указывает на четкое «разделение обязанностей» в химической защите растения — возможно, в ответ на атаки разных видов фитофагов.
А что же долгоносики? Сравнение геномов корневого и стеблевого долгоносиков показало, что их хромосомы похожи, но не идентичны. В них сохранилось много общих «синтенных блоков» — участков ДНК, расположенных в одинаковом порядке, — однако за миллионы лет произошло немало перестроек. Например, у корневого долгоносика девять хромосом соответствуют всего одной хромосоме у стеблевого. Нашлись и случаи слияния и расщепления хромосом: так, каждая из хромосом Ch11, Ch16 и Ch18 у корневого вида в геноме стеблевого «разошлись» на две отдельные, а Ch15 корневого вида оказалась составлена из кусочков Ch15 и Ch16 стеблевого. Были и обратные ситуации, когда у корневого долгоносика одна хромосома получалась из слияния двух у стеблевого (например, Ch16 — результат объединения Ch17 и части Ch16). В целом картина указывает на то, что когда-то у предков этих насекомых был общий набор хромосом, но после разделения видов геномы постепенно «перекроились» — возможно, в ответ на освоение разных частей айланта: корней и стволов. При этом основная масса генов все еще остается близкородственной — филогенетический анализ показал, что пары генов у двух видов группируются вместе, что говорит либо о сравнительно недавнем разделении (около 39 млн лет назад), либо о сходном отборе, действующем на оба вида.

Рис. 4. Схема соответствия между хромосомами двух видов долгоносиков —Eucryptorrhynchus scrobiculatus (верхний ряд) и E. brandti (нижний ряд). Цветные полосы показывают, какие участки хромосом у одного вида соответствуют участкам у другого. Видно, что после разделения видов произошло много перестроек: слияний, расщеплений и перемещений участков. Рисунок из обсуждаемой статьи
Особое внимание ученые уделили генам, связанным с детоксикацией растительных ядов. В целом, набор таких генов у обоих видов оказался похож, за исключением одного заметного отличия — семейства GST. Эти гены кодируют ферменты, которые обезвреживают токсичные соединения. И у корневого, и у стеблевого долгоносика GST-гены образуют компактные кластеры на хромосоме 15: у первого — восемь генов в кластере (из 36 всего), у второго — шесть (из 39). Остальные GST-гены распределены по разным хромосомам. Интересно, что гены внутри этих кластеров у разных видов оказались генетически ближе друг к другу, чем к другим GST-генам в пределах одного вида. Это намекает на то, что кластеры появились до разделения видов и затем эволюционировали параллельно. Некоторые из этих генов работают особенно активно в антеннах, вероятно, помогая разрушать молекулы запахов, другие — в средней кишке, где идет переваривание и обезвреживание растительных ядов. Дупликации генов, участвующих в детоксикации — явление не уникальное, оно часто встречается у фитофагов, которые питаются растениями с сильной химической защитой. Похоже, что и в случае долгоносиков именно дублирование GST-генов, вместе с их тканеспецифичной активностью, стало ключевым шагом в адаптации к жизни на айланте. Хотя, как отмечают авторы, это еще предстоит подтвердить экспериментально.
По сравнению с другими жуками, питающимися на разных растениях, набор генов, отвечающих за запахи, у этих строго специализированных монофагов оказался скромным, что согласуется с наблюдаемой в мире насекомых тенденцией: чем шире спектр кормовых растений, тем больше генов хеморецепторов. Обычно именно расширение этих семейств помогает насекомым быстро подстраиваться под новые условия. Однако в геномах двух долгоносиков заметное «разрастание» оказалось только в семействе SNMP (гены мембранных белков сенсорных нейронов, которые играют ключевую роль в обонянии насекомых). У корневого вида нашли 11 таких генов, у стеблевого — 12. Среди них были представители подсемейств SNMP1 и SNMP2, но неожиданно оказалось, что девять генов относятся к ранее не описанному у жуков подсемейству SNMP5. Интересно, что до этого SNMP5 встречалось в основном у тараканов, трипсов и полужесткокрылых — насекомых, которые, как и взрослые долгоносики, питаются флоэмой растений. Это навело исследователей на мысль, что общая эволюционная история и сходный тип питания могли способствовать сохранению SNMP5 у этих групп, тогда как у большинства жуков оно было утрачено.
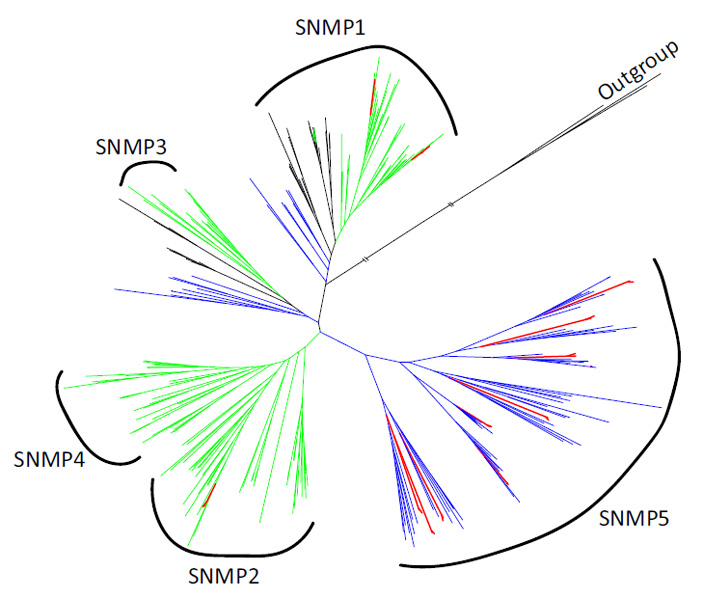
Рис. 5. Филогенетическое дерево генов сенсорных нейронных мембранных белков (SNMP) у двух видов долгоносиков (оба вида показаны красным) и других насекомых. Зеленым показаны другие жесткокрылые, синим и черным — представители других отрядов (включая Diptera, Hymenoptera, Lepidoptera — черный, Hemiptera, Thysanoptera, Blattodea — синий). Дерево разделено на пять основных клад (SNMP1–SNMP5); видно, что у долгоносиков обнаружено расширение в подсемействе SNMP5, ранее редко встречавшемся у жуков. Рисунок из обсуждаемой статьи
Изучение активности генов показало, что гены семейства SNMP5 у обоих долгоносиков «включены» на всех стадиях развития и во всех тканях, причём их уровень экспрессии выше, чем у семейств SNMP1 и SNMP2. Специфической привязки к полу или определенным органам у них не обнаружили. Зато у SNMP1 картина была другой: единственный ген этого типа (EscrSNMP1) у корневого долгоносика работал в антеннах самцов и самок почти в сто раз активнее, чем в других тканях, что указывает на его возможную роль в восприятии половых феромонов. У стеблевого вида аналогичную функцию выполняли два гена — EbraSNMP1a и EbraSNMP1b, которые, по сути, являются разными частями одного «двойного» гена у корневого собрата.
Чтобы понять, как эволюция влияла на эти гены, исследователи проверили их на признаки отбора. В SNMP у корневого долгоносика нашли девять участков, находящихся под действием положительного (диверсифицирующего) отбора, у стеблевого — восемь. Для сравнения, в другом важном семействе — GST — таких участков было 14 и 8, соответственно. Аналогичный анализ провели и для вовлеченных в биосинтез квассиноидов генов айланта, в которых также были обнаружены участки положительного отбора. При этом подавляющее большинство позиций в этих генах у насекомых и растения находилось под действием сильного отрицательного (очищающего) отбора, что говорит о важности их сохранения в стабильном виде.
В совокупности результаты показывают, что айлант и его специализированные фитофаги на протяжении миллионов лет находятся в состоянии эволюционной «гонки вооружений». Расширение основных генов, вовлеченных в биосинтез квассиноидов у айланта, было связано с тандемными дупликациями и дупликацией всего генома, тогда как расширение генов детоксифицирующих ферментов GST у долгоносиков происходило за счет тандемных дупликаций, а генов хемосенсорных белков SNMP — за счет появления новых генов. Это исследование указывает на то, что как растения, так и насекомые-фитофаги перестраивали свои геномы путем расширения семейств генов, при этом у растения произошло также удвоение всего генома, а у двух видов долгоносиков — формирование новых генов. Эти изменения, вероятно, отражают гонку вооружений между защитой и контрзащитой, демонстрируя, как трофические взаимодействия могут направлять эволюцию геномов.
Источник: Wei Song, Li-Jun Cao, Jin-Cui Chen, Wen-Juan Guo, Hui-Juan Li, Xue-Wen Sun, Ary Anthony Hoffmann, Jun-Bao Wen, Shu-Jun Wei. Genomes of Two Monophagous Weevils and Their Host Plant Provide Insights Into Evolution of Plant Defence and Insect Counter‐Defence // Molecular Ecology Resources. 2025. DOI: 10.1111/1755-0998.70009.
Елена Устинова


Но это, мне кажется, пока глубоко в тему не залезли...
Прочий опыт показывает, что чем глубже - тем сложнее, и конца-края нету)
Кстати говоря, камуфляж у жуков сих бросился в глаза. Что ж они так по-военному покрашены? Прям снайперы будто - фиг разглядишь на фоне естественной среды. От кого прячутся? И почему бы в анализе генома такую серьёзную работу над незаметностью не учесть/оценить?




Рис. 1. Айлант высочайший (слева) и специализирующиесф на нем долгоносики Eucryptorrhynchus scrobiculatus (справа вверху) и E. brandti (справа внизу). Фото айланта с сайта treeseed.ru. Фото долгоносиков с сайта inaturalist.org