Панцирное бесчелюстное туцзяаспис проливает свет на происхождение конечностей позвоночных

Китайские и британские палеонтологи описали из раннесилурийских отложений Китая представителя панцирных бесчелюстных из класса Galeaspida с двумя длинными плавниковыми складками, тянущимися от головы до хвоста по краям плоского брюха. Гидродинамические расчеты показали, что эти складки создавали подъемную силу при движении вперед. Возможно, это помогало древнему рыбообразному спасаться от хищников. Находка неплохо согласуется с классической гипотезой, согласно которой парные конечности челюстноротых позвоночных (грудные и брюшные плавники, ставшие у тетрапод передними и задними ногами) сформировались в результате фрагментации исходно цельных плавниковых складок. При этом первыми, по-видимому, обособились грудные плавники, и лишь потом оставшаяся часть плавниковой складки компактизовалась и превратилась в брюшные плавники. Промежуточное состояние — с грудными плавниками и плавниковыми складками позади них — характерно для другой группы панцирных бесчелюстных, Osteostraci, которая на эволюционном дереве занимает промежуточное положение между Galeaspida и челюстноротыми.
Когда и каким образом позвоночные обзавелись парными конечностями — грудными и брюшными плавниками, которые позже в одной из линий (у тетрапод) превратились в ноги с пальцами? Точного ответа ученые до сих пор не знают. У современных бесчелюстных (миног и миксин) парных конечностей нет и, видимо, никогда не было. У челюстноротых (Gnathostomata) они есть, причем как у современных групп (хрящевых и костных рыб, включая тетрапод), так и у вымерших плакодерм. По-видимому, парные плавники впервые появились на каком-то этапе эволюции остракодерм — вымерших бесчелюстных рыбообразных, занимающих на эволюционном дереве промежуточное положение между современными бесчелюстными и челюстноротыми. Проблема в том, что от остракодерм в ископаемом состоянии обычно сохраняются только твердые элементы панциря: головные щиты, отдельные костные пластины или чешуи. По более нежным частям тела, таким как плавники, ископаемая летопись остракодерм предоставляет довольно скудные данные.
Статья китайских и британских палеонтологов, опубликованная в конце сентября в журнале Nature, существенно дополняет эти данные и проливает новый свет на проблему происхождения парных конечностей позвоночных. Исследователи обнаружили в раннесилурийских (возраст около 436 млн лет) отложениях Южного Китая (провинция Хунань) три превосходно сохранившихся, почти не деформированных экземпляра нового вида мелких (до 7 см) остракодерм из группы Galeaspida. Новооткрытый представитель панцирных бесчелюстных (по-русски не принято называть остракодерм «рыбами», хотя в англоязычной литературе термин «jawless fish» вполне допустим) получил название Tujiaaspis vividus (Tujia — туцзя, народность, проживающая в тех краях, aspis — щит, vividus — одухотворенный, полный жизни).
Реконструкции T. vividus показаны на рис. 1 и 4, типовой экземпляр с прорисовкой — на рис. 2. По строению костного головного щита (той части тела, которая у остракодерм сохраняется лучше всего) туцзяаспис — типичный представитель галеаспид. При этом у него есть специфические особенности в строении сенсорных каналов, пронизывающих щит (ifc, ldc, ltc на рис. 2), что позволило выделить его в новый род и вид.

Рис. 2. Голотип Tujiaaspis vividus. Слева — фотография, справа — прорисовка. Длина масштабного отрезка — 4 мм, длина животного — около 7 см. Хорошо видна пара «плавниковых складок» — длинных вентролатеральных плавников (vlf). Рисунок из обсуждаемой статьи в Nature
Самое важное в новых находках то, что у туцзяасписа отлично сохранились плавники: три спинных (у голотипа виден только один из них, df3 на рис. 2), хвостовой (двухлопастной, причем нижняя лопасть (al) является продолжением оси тела; такой плавник называют гипоцеркальным). И, наконец, главное: пара длинных вентролатеральных плавников (или плавниковых складок), которые тянутся по бокам уплощенного брюха от жаберной области до самого хвоста (vlf на рис. 2). Вентролатеральные плавники поддерживались многочисленными V-образными скелетными элементами и вдобавок были покрыты мелкими чешуйками.

Рис. 3. Реконструкция Cephalaspis salweyi, раннедевонского представителя Osteostraci (вид снизу). Видны парные грудные плавники и «вентролатеральные гребни», тянущиеся по краям плоского брюха. Фото с сайта blogs.ucl.ac.uk
Морфологические структуры, напоминающие вентролатеральные плавники туцзяасписа, ранее были описаны у некоторых представителей других групп остракодерм. В частности, у Anaspida тоже встречаются длинные вентролатеральные плавниковые складки. У Thelodonti — раздельные грудные и брюшные парные плавниковые складки (лишенные, в отличие от настоящих плавников, внутреннего скелета). У некоторых Osteostraci (группа остракодерм, наиболее близкая к челюстноротым) есть очень похожие «вентролатеральные гребни», но они не доходят спереди до жаберной области, и к ним в передней части туловища прилагаются уже дифференцированные грудные плавники (рис. 3). Что касается галеаспид, которые считаются сестринской группой по отношению к группе [челюстноротые + Osteostraci], то у них ситуация с плавниками (как парными, так и непарными) до сих пор оставалась неясной. Хорошо изучены у галеаспид фактически были только головные щиты.
Авторы изучили гидродинамические свойства виртуальных трехмерных моделей туцзяасписа (рис. 4) и пришли к выводу, что вентролатеральные плавники сильно влияют на эти свойства. Они создают значительную подъемную силу при движении вперед. По-видимому, вентролатеральные плавниковые складки позволяли туцзяаспису быстро всплывать надо дном. Возможно, это помогало ему удирать от хищников, таких как крупные ракоскорпионы (рис. 1). Вентролатеральные гребни Osteostraci, скорее всего, выполняли ту же функцию.

Рис. 4. Виртуальная реконструкция туцзяасписа, которую использовали для изучения его гидродинамических свойств. Рисунок из обсуждаемой статьи в Nature
Авторы предполагают, что наблюдающееся у остракодерм разнообразие парных выростов и складок по бокам тела — независимо от того, гомологичны ли они парным плавникам челюстноротых, — отражает попытки эволюции обеспечить этих тяжеловесных, неповоротливых бесчелюстных хоть какой-то маневренностью и способностью быстро всплывать при отсутствии нормальных грудных и брюшных плавников с их внутренним скелетом и специализированными мышцами. Скорее всего, такие складки возникали несколько раз независимо в разных группах остракодерм. Однако вентролатеральные плавниковые складки Galeaspida и Osteostraci авторы считают гомологичными образованиями, потому что они очень похожи как по структуре, так и по расположению. Впоследствии эти складки превратились в раздельные грудные и брюшные плавники челюстноротых.
По мнению авторов, морфологические изменения в ряду Galeaspida — Osteostraci — Gnathostomata отражают ход эволюции парных конечностей: сначала неразделенные, длинные вентролатеральные плавниковые складки от жабер до хвоста (Galeaspida), потом их передние части обособляются в грудные плавники (Osteostraci), и, наконец, оставшаяся часть вентролатеральных складок компактизуется и превращается в брюшные плавники (Gnathostomata). При этом не исключено, что Galeaspida унаследовали свои вентролатеральные плавниковые складки от более древних бесчелюстных.
Вырисовывающаяся картина очень похожа на старую, классическую идею о том, что парные плавники рыб развились из некогда единой боковой плавниковой складки. Эту гипотезу развивали, в частности, А. Н. Северцов и И. И. Шмальгаузен. Но есть нюанс: классическая гипотеза предполагала более или менее одновременное формирование обеих пар конечностей путем фрагментации исходно единой складки, а новая схема предполагает, что сначала в передней части туловища обособились грудные плавники. На этом этапе остаток плавниковой складки продолжал обеспечивать пассивное создание подъемной силы (Osteostraci). Потом, уже у первых челюстноротых, этот остаток съежился и дал начало брюшным плавникам. Новая схема в целом неплохо соответствует палеонтологическим данным (рис. 5). К этому стоит добавить, что у древних челюстноротых, вероятно, еще сохранялась потенциальная возможность формирования плавников вдоль всей линии, соединяющей грудные плавники с брюшными, как видно из морфологии акантод с их рядами парных плавниковых шипов между грудными и брюшными плавниками.
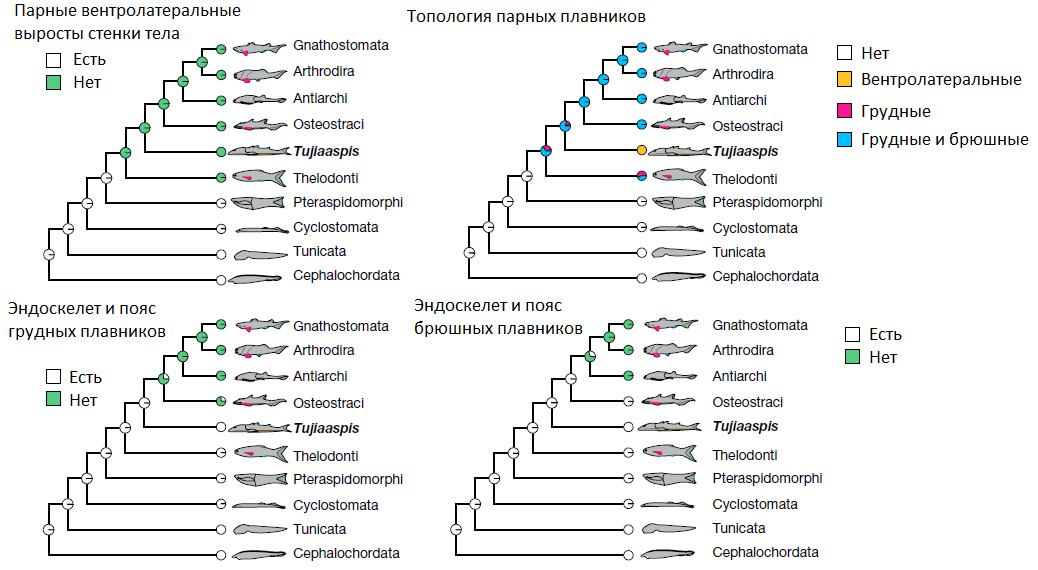
Рис. 5. Схема эволюции парных конечностей с учетом новых данных по галеаспидам (Tujiaaspis). Цветные кружочки отражают вероятности того или иного состояния признака у предковых форм. Arthrodira и Antiarchi — челюстноротые панцирные рыбы или плакодермы, Gnathostomata — «настоящие» челюстноротые (за вычетом плакодерм; в тексте новости под этим названием подразумеваются все челюстноротые), Cyclostomata — современные бесчелюстные (круглоротые), Tunicata — оболочники, Cephalochordata — головохордовые (ланцетник), остальные группы — различные остракодермы. Рисунок из обсуждаемой статьи в Nature
Морфология туцзяасписа, возможно, проясняет еще один спорный момент в происхождении парных плавников. Давно замечено, что парные плавники сходны с непарными (спинными, хвостовым) по многим деталям своего строения и развития. Поскольку непарные плавники в ходе эволюции появились раньше парных, высказывалось предположение, что происхождение парных плавников включало коопцию (то есть повторное использование для новой задачи) регуляторных генных каскадов, изначально служивших для регуляции развития непарных плавников. Это подтверждается молекулярно-генетическими данными: некоторые гены (в том числе из групп Hox и Tbx), сигнальные молекулы (SHH, FGF) и регуляторные участки генома (энхансеры) задействованы в развитии как парных, так и непарных плавников. Найденные экземпляры туцзяасписа сохранились настолько хорошо, что на них видно, откуда в принципе могло пойти это глубинное cходство («глубокая гомология», см. Deep homology) парных и непарных плавников. Дело в том, что вентролатеральные плавниковые складки туцзяасписа на заднем конце тела фактически сливаются с хвостовым плавником и являются как бы его раздвоенным продолжением. Это намекает на возможное происхождение плавниковых складок путем расширения в переднем направлении области экспрессии тех генов, которые изначально контролировали развитие хвостового плавника. Или, возможно, сначала был непарный длинный плавник вдоль середины брюха, который потом раздвоился спереди, сохраняя поначалу связь с хвостовым плавником, который остался непарным.
Возможно, лишь немного преувеличивая, можно сказать, что до публикации обсуждаемой статьи попытки разобраться в происхождении парных плавников, роясь в литературе по ископаемым рыбообразным, мало что могли дать, кроме головной боли и ощущения полной неясности. Теперь же появилось хоть что-то похожее на внятную эволюционную гипотезу с весомыми палеонтологическими основаниями.
Источник: Zhikun Gai, Qiang Li, Humberto G. Ferrón, Joseph N. Keating, Junqing Wang, Philip C. J. Donoghue & Min Zhu. Galeaspid anatomy and the origin of vertebrate paired appendages // Nature. 2022. DOI: 10.1038/s41586-022-04897-6.
См. также:
1) В силурийских отложениях Китая найден древнейший позвоночный хищник, «Элементы», 16.06.2014.
2) Открыто ископаемое, сочетающее признаки панцирных и костных рыб, «Элементы», 02.10.2013.


А у Маркса таинственные "производительные силы", действующие во вселенной. У Ницше - воля к власти (жизни) и круговращение ее в природе. У Фрейда либидо итд. Каждый верит в свою версию невидимых сил. Да, все они, в отличии от идеалистической версии (творца) - тупые (действуют методом подбора и ошибок) и онтологически не существуют, но некие хитроумные люди, типа Дорвина вскрыли их работу.
Новости: Палеонтология












Рис. 1. Художественная реконструкция Tujiaaspis vividus. На рисунке также присутствует ракоскорпион (эти крупные членистоногие могли охотиться на мелких панцирных бесчелюстных, подобных туцзяаспису) и стайка конодонтоносителей. Рисунок Qiuyang Zheng из обсуждаемой статьи в Nature