Судя по строению вестибулярного аппарата, пермские антеозавры были активными и ловкими хищниками

Длительное время считалось, что пермские терапсиды — прямые предки современных млекопитающих — были грузными, медлительными и неуклюжими созданиями, с трудом передвигавшимися на своих полувыпрямленных конечностях и способных обеспечивать себя кислородом только благодаря его повышенному содержанию в атмосфере, а никак не эффективной системе дыхания. Но недавнее томографическое исследование черепа одного из самых крупных хищных диноцефалов — южноафриканского антеозавра (Anteosaurus magnificus) — показало, что при жизни этот шестисоткилограммовый монстр был достаточно подвижным и вполне мог активно преследовать свою жертву, как это делают современные плотоядные млекопитающие.
Диноцефалы доминировали на Земле в середине пермского периода (270–260 миллионов лет назад), представляя собой многочисленную и разнообразную группу четвероногих животных, освоивших, судя по имеющимся находкам чуть ли не всю Пангею. Предковой формой диноцефалов, скорее всего, являются биармозухи (см. Биармозух и очёрская фауна), примитивные хищные терапсиды, обнаруженные в Приуралье, тогда как все более поздние группы этих животных, скорее всего, зародились на юге, и лишь потом часть из них мигрировала на север. Предполагается, что к примитивным диноцефалам могут относиться и эстемменозухи (см. картинку дня Коронованный «крокодил»), но это неточно, ибо некоторые исследователи сближают эстемменозуха с горгонопсами. Традиционно к диноцефалам относят представителей двух инфраотрядов: преимущественно растительноядных тапиноцефалов (Tapinocephalia) и плотоядных антеозавров (Anteosauria).
Тапиноцефалы в основном были представлены крупными формами (например, знаменитый мосхопс достигал 2,7 м в длину и весил полторы тонны, а давший имя всей группе тапиноцефал был еще крупнее) с очень характерным внешним видом: коротким хвостом, длинными передними конечностями (из-за этого плечи диноцефала располагались значительно выше крестца) и массивной головой с выраженным пахиостозом (см. Pachyostosis) лобных костей. Точное назначение этих костных утолщений пока что остается неясным: зарубежные исследователи предполагают, что они помогали при внутривидовых стычках (см. E. Snively, A. Cox, 2007. Structural mechanics of Pachycephalosaur crania permitted head-butting behavior), а вот отечественные палеонтологи чаще придерживаются версии, что пахиостозные нарастания костей диноцефалов, пронизанные крупными кровеносными сосудами и не защищенные никакими роговыми образованиями, служили для терморегуляции и помогали этим гигантским животным не перегреваться на солнце (M. Ivakhnenko, 2008. Cranial morphology and evolution of Permian Dinomorpha (Eotherapsida) of eastern Europe).
Антеозавры считаются более примитивными животными по сравнению с тапиноцефалами, поскольку и по образу жизни, и по общим чертам строения скелета они больше схожи с примитивными диноцефалами, чем со своими растительноядными современниками. У них были более тонкие кости черепа (см. C. Kammerer, 2009. Systematics of the Anteosauria (Therapsida: Dinocephalia), L. Boonstra, 1969. The fauna of the Tapinocephalus zone (Beaufort beds of the Karoo)), а пахиостозные утолщения, предположительно, развились для поглощения нагрузки, создаваемой мощной приводящей мускулатурой во время нанесения укусов, так что для них «бодание» между самцами представляется еще менее вероятным. Разобраться с ролью этих костных образований у антеозавров при помощи современных методов попыталась группа ученых из Южной Африки, Швеции и Великобритании. Они провели рентгеновскую микрокомпьютерную томографию черепа молодого антеозавра BP/1/7074, позволившую в буквальном смысле заглянуть внутрь его головы. Ранее такие исследования проводились только для растительноядных тапиноцефалов (поскольку строение их эволюционно продвинутых черепов больше интересовало ученых), тогда как нейроанатомия хищных диноцефалов оставалась практически неизученной.

Рис. 2. Два крупных диноцефала — плотоядный антеозавр (слева) и растительноядный мосхопс (справа). Рисунок © John Sibbick с сайта k.sina.com.cn
Первой интересной особенностью, выявленной при томографии черепа антеозавра, было строение его внутреннего уха: по сравнению с мосхопсом, у антеозавра оказались гораздо более длинные и сильнее изогнутые полукружные каналы, что предполагает отлично развитое чувство равновесия, характерное для достаточно подвижного животного. В целом размер внутреннего уха антеозавра, относительно размеров тела, больше, чем у любого другого известного пермского терапсида, как растительноядного, так и хищного: по мнению исследователей это означает, что антеозавр мог быстро двигать глазами, головой и шеей, а значит — ловко выслеживать и настигать свою жертву, опережая других плотоядных. Но поскольку череп BP/1/7074 принадлежит молодому животному, нельзя сказать с уверенностью, сохранялись ли подобные пропорции в строении внутреннего уха при достижении зрелости: к сожалению, онтогенетические исследования терапсид крайне редки. Единственные доступные на сегодняшний день данные — это исследование строения двух черепов разновозрастных цинозавров (Cynosaurus), терапсид-цинодонтов, наиболее близких к млекопитающим: судя по относительным размерам и строению внутреннего уха, как минимум у части пермских терапсид внутреннее ухо достигало анатомической зрелости в сравнительно раннем возрасте, как это характерно для современных млекопитающих (J. Benoit et al., 2017. The bony labyrinth of late Permian Biarmosuchia: palaeobiology and diversity in non-mammalian Therapsida).
Гипотеза о достаточно подвижном образе жизни подтверждается наличием глубокой и широкой флоккулярной ямки, в которой при жизни располагался клочок (flocculus) мозжечка — наиболее древняя часть этого раздела мозга, являющаяся важнейшей частью вестибулярного аппарата. Для сравнения, у мосхопса эта ямка была намного меньше и не такой глубокой, так что при жизни мосхопс был определенно медлительнее своего хищного родственника, чей образ жизни теперь представляется вовсе не таким малоподвижным, как его ранее описывали некоторые палеонтологи (см., например, G. King, 1988. Encyclopedia of Paleoherpetology и M. Ivakhnenko, 2008. Cranial morphology and evolution of Permian Dinomorpha (Eotherapsida) of eastern Europe).
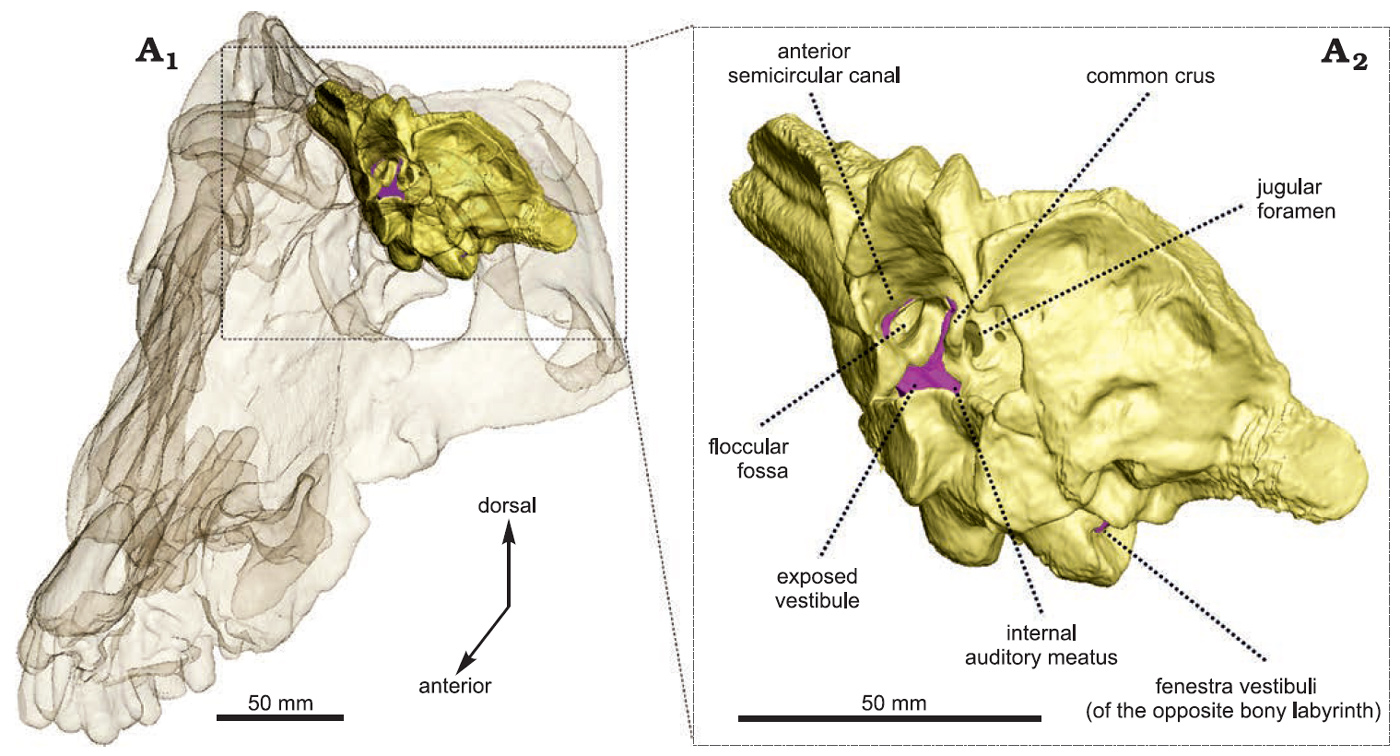
Рис. 3. Строение затылочной кости антеозавра (выделена желтым) и костного лабиринта, включающего внутреннее ухо (выделен фиолетовым). Anterior semicircular canal — передний полукружный канал, common crus — общая ножка, jugular foramen — яремное отверстие, floccular fossa — флоккулярная ямка, exposed vestibule — преддверие уха, internal auditory meatus — внутренний слуховой проход, fenestra vestibule — окно преддверия (противоположного уха). Рисунок из обсуждаемой статьи в Acta Palaeontologica Polonica
Помимо строения вестибулярного аппарата исследователи также обратили внимание на ориентацию мозговой полости антеозавра по отношению к остальному черепу. У антеозавра длинная ось мозговой полости практически параллельна оси черепа (условной линии от затылочного мыщелка до кончика морды), в то время как у мосхопса при горизонтальной ориентации мозговой полости кончик морды направлен круто вниз. Дополнительно такую ориентацию можно подтвердить сравнением угла между плоскостью бокового полукружного канала и длинной осью черепа: у антеозавра этот угол составляет 25°, а у мосхопса — 65°. Авторы предполагают, что в ходе эволюции у тапиноцефалов произошла значительная реорганизация исходного строения черепа, которая, вероятно, была вызвана адаптацией к использованию головы в качестве оружия или к специфическому способу питания. У современных млекопитающих такая переориентация черепа относительно длинной оси мозговой полости называется клиноринхией (см. Primate Cranial Diversity) и обычно интерпретируется как адаптация к травоядности и боданию у копытных, а у приматов — как следствие увеличения мозга. Поскольку у тапиноцефалов мозг относительно невелик, предполагается, что наклон их черепа сформировался по тем же причинам, что и у современных овец или коз, тогда как у антеозавров ничего подобного не наблюдается. Возможно, если хищные диноцефалы и устраивали межсамцовые баталии, то они вели себя скорее не как современные благородные олени, а как кабарги, которые используют свои увеличенные верхние клыки и для демонстрации, и в качестве оружия во время конфликтов.
Все эти соображения с учетом данных предыдущих изысканий позволили авторам обсуждаемой рабты предположить, что в ходе эволюции примитивные диноцефалы, у которых пахиостозные утолщения черепа в основном служили для укрепления костей и снижения нагрузки при укусах, постепенно начали преобразовываться в более прогрессивных тапиноцефалов, у которых пахиостоз внушительно разросся (у некоторых видов толщина черепной коробки в области лба достигала 30 сантиметров, см. M. Day et al., 2015. Youngest dinocephalian fossils extend the Tapinocephalus Zone, Karoo Basin, South Africa) и начал выполнять социальные функции.

Рис. 4. Расположение мозговой полости антеозавра (А) и мосхопса (B) по отношению к основному черепу. Pineal (parietal) foramen — теменной глаз, occipital condyle — затылочный мыщелок, fighting surface of the skull — пахиостоз черепа, согласно предположению авторов, используемый при бодании, floccular fossa — флоккулярная ямка. Рисунок из обсуждаемой статьи в Acta Palaeontologica Polonica
Таким образом, новые данные (положение мозговой полости антеозавра относительно черепа, строение его внутреннего уха и размер флоккулярной ямки) подтверждают, что антеозавры не были неуклюжими амфибиотическими хищниками, а отличались значительной подвижностью и ловкостью по меркам пермских терапсид — что, учитывая последние данные изотопного анализа их костей (K. Rey et al., 2020. Oxygen isotopes and ecological inferences of Permian (Guadalupian) tetrapods from the main Karoo Basin of South Africa), позволяет представить этих животных аналогами современных медведей, а не крокодилов, как считалось ранее. Также изучение внутренней анатомии черепа антеозавра говорит в пользу гипотезы, что в то время как тапиноцефалы были стадными животными с довольно развитым социальным поведением (что косвенно подтверждается находками групп тапиноцефалов из 5–12 особей, B. Rubidge et al., 2019. The postcranial skeleton of the basal tapinocephalid dinocephalian Tapinocaninus pamelae (Synapsida: Therapsida) from the South African Karoo Supergroup), антеозавры отличались более примитивным строением черепа и, скорее всего, вели образ жизни хищников-одиночек, как это характерно и для многих современных плотоядных млекопитающих.
Источник: J. Benoit, A. Kruger, S. Jirah, V. Fernandez, B. S. Rubidge. Palaeoneurology and palaeobiology of the dinocephalian therapsid Anteosaurus magnificus // Acta Palaeontologica Polonica. 2021. DOI: 10.4202/app.00800.2020.
Анна Новиковская
-
Научно-художественная реконструкция прижизненного облика по скелетам трудная конечно штука. Кажется, здесь же на Элементах кто-то приводил в пример реконструкцию бегемота. Если исходить только из скелета, то получается ужасный монстр. В то время как в реальности внешне довольно милое и дружелюбное создание. Дело в деталях, например, фактура кожи, наличие шерсти, губ, кожных складок, количество подкожного жира, цвет и т.п.
Здесь на рис.2, например, кожа нарисована скорее как у рептилий. В то время как у синапсид кожа унаследована от земноводных, должна быть гладкая и эластичная.-
Тут, быть может, проблема в том, что на рисунке 2 представлена довольно старая реконструкция из тех времен, когда синапсид все еще на полном основании считали рептилиями и реконструировали как ящериц с собачьими зубами. :) Хотя, честно говоря, у некоторых современных реконструкторов проблемы те же...
-
Здесь на рис.2, например, кожа нарисована скорее как у рептилий. В то время как у синапсид кожа унаследована от земноводных, должна быть гладкая и эластичная.
Это заблуждение. Наличие чешуи было общим признаком для древних амниот - базальных синапсид и рептилий. У синапсид впоследствии она преобразовалась в волосяной покров, у рептилий (а именно у эурептилий, что там с парарептилиями - точно не известно) стала кератиновой, что лишило их кожных желез.
Среди базальных синапсид чешуя найдена у офиакодонтов, диметродонов (походила на крокодилью - крупные бляшки) и у варанопид (вылитые ящерицы).
С отпечатками кожи базальных терапсид (биармозухий) повезло меньше, какая у них была кожа точно не известно. Они ближе всего к диметрадонам, так что могли иметь чешую как у них.
Есть упоминания, что у эстеменозухов (дейноцефалы) в коже были какие-то мелкие окостенения, но без сплошного чешуйчатого покрова.
Чешуя у офиакодонтов
https://www.researchgate.net/publication/292461005_Pennsylvanian_pelycosaurs_from_Linton_Ohio_and_Nyrany_Czechoslovakia
У диметродонов
https://www.researchgate.net/publication/233780818_A_Supposed_Eupelycosaur_Body_Impression_from_the_Early_Permian_of_the _Intra-Sudetic_Basin_Poland
У варанопид
https://www.researchgate.net/publication/323782950_First_arboreal_'pelycosaurs'_Synapsida_Varanopidae_from_the_early_Per mian_Chemnitz_Fossil_Lagerstatte_SE_Germany_with_a_review_of _varanopid_phylogeny
Там же смотри чешую (Fig. 26 c,d) на веках темноспондильных амфибий, такая же как у варанопид, описанных в статье.
И ещё чешуя у темноспондильных амфибий
https://www.researchgate.net/publication/26340018_On_the_squamation_of_Australerpeton_cosgriffi_Barberena_a_temnospondyl _amphibian_from_the_Upper_Permian_of_Brazil -
Позволю не согласиться насчет эстемменозухид. Во всяком случае, и Чудинов, и Ивахненко писали, что, судя по сохранившимся отпечаткам кожи, шкура у терапсид была мягкой, без явно выраженного фолидоза, а кальцинизированные линзочки, сохранившиеся в ней - это остатки каких-то кожных желез типа потовых. Максимум "чешуи", на которую терапсиды могли рассчитывать - это тонкие кератиновые чешуйки, как на крысином хвосте.
Что касается варанопид, то они же теперь выведены из состава синапсид, нэ? Так что, если чешуя и была, то только у пеликозавров.
-
-
-
Интересно, догадались бы мы, что зайцы длинноухи, будь они ископаемыми? (А ежи колючи?) Кстати, а вдруг терапсиды были с длинными ушами? Сейчас в пэйнте пририсовал уши антеозавру (рис. 1), прикольно получилось. Хотя, если подумать, у зайца должны быть кости, к которым прикрепляются мышцы, двигающие ушами, и это можно было бы понять по костям. И если бы у терапсид были длинные уши, то у них были бы аналогичные костные образования, и их бы уже давно заметили.


Да... Странные были времена.
А, конечно, факт, что можно окаменелость рентгеном изучать - всегда поражал. Камень, казалось бы, чего там разглядишь?
И коль скоро это такой действенный способ - сегодня рентген не слишком редок и дорог, в каждой поликлинике стоит, и очереди на стоят к ним - отчего так мало применяется археологами?
Анапсиды, диапсиды, синапсиды
-
 13.04.2026В бассейне Кару на юге Африки впервые обнаружено яйцо синапсидыАнна Новиковская • Новости науки
13.04.2026В бассейне Кару на юге Африки впервые обнаружено яйцо синапсидыАнна Новиковская • Новости науки -
 06.12.2022Растительноядные экзаэретодоны баловали детей более калорийной животной пищейАнна Новиковская • Новости науки
06.12.2022Растительноядные экзаэретодоны баловали детей более калорийной животной пищейАнна Новиковская • Новости науки
-
 13.10.2022Нора дииктодонаАнна Новиковская • Картинки дня
13.10.2022Нора дииктодонаАнна Новиковская • Картинки дня
-
 05.09.2022Найдены мумии листрозавров, живших в начале триасового периодаАнна Новиковская • Новости науки
05.09.2022Найдены мумии листрозавров, живших в начале триасового периодаАнна Новиковская • Новости науки
-
 29.12.2021Проколофон из кернаАнтон Ульяхин • Картинки дня
29.12.2021Проколофон из кернаАнтон Ульяхин • Картинки дня
-
 02.04.2021Судя по строению вестибулярного аппарата, пермские антеозавры были активными и ловкими хищникамиАнна Новиковская • Новости науки
02.04.2021Судя по строению вестибулярного аппарата, пермские антеозавры были активными и ловкими хищникамиАнна Новиковская • Новости науки
-
 11.12.2020«До начала зверей». Главы из книгиАнна Новиковская • Книжный клуб • Главы
11.12.2020«До начала зверей». Главы из книгиАнна Новиковская • Книжный клуб • Главы
-
 23.11.2020Мегавайтсия, ядовитый медведеящерАнна Новиковская • Картинки дня
23.11.2020Мегавайтсия, ядовитый медведеящерАнна Новиковская • Картинки дня
-
 17.11.2020«Непонятные ящерицы»Анна Новиковская • Картинки дня
17.11.2020«Непонятные ящерицы»Анна Новиковская • Картинки дня
-
 09.11.2020ЛисовицияАнна Новиковская • Картинки дня
09.11.2020ЛисовицияАнна Новиковская • Картинки дня
Последние новости













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Рис. 1. Реконструкция внешнего вида антеозавра (Anteosaurus magnificus). Рисунок © Виталий Мельник с сайта malvit.artstation.com