Новое применение для древней регуляторной системы: длина крыльев у рисовой цикадки регулируется инсулиновым сигнальным каскадом

Китайские ученые расшифровали молекулярный механизм, благодаря которому бурая рисовая цикадка — опаснейший вредитель рисовых плантаций — развивается по одному из двух альтернативных путей, становясь либо длиннокрылой мигрирующей формой, либо короткокрылой, нелетающей и быстро размножающейся. Как выяснилось, ключевую роль в регуляции развития крыльев играет инсулиновый каскад — древняя сигнальная система, которая у многих животных, включая насекомых и млекопитающих, отвечает за адаптацию организма к колебаниям количества доступной пищи.
Полифенизмом (см.: Polyphenism) называют способность одного и того же генотипа обеспечивать развитие двух или более дискретных фенотипов, причем выбор пути развития зависит от внешних условий (см. ссылки в конце новости). Типичный пример полифенизма — касты у общественных перепончатокрылых насекомых.
Для многих насекомых характерен так называемый крыловой полифенизм. Это значит, что в одних условиях насекомое отращивает крылья и совершает миграции в поисках более благоприятных мест для жизни, а в других из точно такой же личинки развивается взрослая особь с недоразвитыми крыльями, нелетающая, но зато способная к ускоренному размножению. Внешние условия, от которых зависит выбор пути развития, могут быть разными, но чаще всего они связаны с состоянием кормовой базы. Пока еды кругом вдоволь, выгоднее никуда не улетать, а побыстрее есть и размножаться. Но когда начинает ощущаться нехватка пищи (например, из-за возросшей численности едоков), более оправданной может стать стратегия мигранта.
Именно такой сезонный крыловой полифенизм характерен для бурой рисовой цикадки Nilaparvata lugens (см. Brown planthopper). Это неприметное насекомое (рис. 1) является чуть ли не главным врагом современного человечества: ведь это опаснейший вредитель риса, составляющего основу питания более половины людей на планете. Рисовая цикадка высасывает соки из стеблей и листьев риса и может приводить к потере значительной части урожая.
И самцы, и самки рисовой цикадки в зависимости от условий могут становиться либо длиннокрылыми мигрирующими формами, либо короткокрылыми оседлыми. На стадии личинки (нимфы) эти формы неразличимы. Однако в результате последней линьки из нимфы вылупляется либо цикадка с двумя парами полностью развитых крыльев, либо с резко укороченными передними крыльями и почти полностью отсутствующими задними.
Зимой рисовые цикадки живут в основном в тропических и субтропических областях Южной Азии. В это время среди них преобладают короткокрылые формы, отличающиеся повышенной плодовитостью. Весной и летом появляется множество длиннокрылых особей, которые мигрируют в северные районы Индии, Китая, в Японию и Корею, где в это время на рисовых полях есть чем поживиться. Осенью стаи длиннокрылых цикадок мигрируют обратно на юг. Потомство, которое они произведут зимой в тропиках, снова будет в основном короткокрылым.
Ранее было показано, что на вероятность развития длиннокрылых и короткокрылых форм влияет плотность популяции и качество пищи. Однако конкретный молекулярный механизм, позволяющий «генетической программе развития» насекомого выбирать один из двух альтернативных путей, не был известен.
Этот механизм удалось частично расшифровать исследователям из Чжэцзянского университета (Ханчжоу, Китай). Поскольку регуляция полифенизма у рисовой цикадки как-то связана с питанием, ученые предположили, что в ней может быть задействован инсулиновый сигнальный каскад (insulin/insulin-like growth factor signalling pathway) — древняя, эволюционно консервативная сигнальная система, которая у многих животных обеспечивает разнообразные приспособительные реакции организма на колебания количества питательных веществ (см.: Insulin signal transduction pathway and regulation of blood glucose).
Чтобы проверить это предположение, авторы стали отключать у цикадок отдельные гены инсулинового каскада, вводя в ткани развивающейся нимфы двухцепочечные молекулы РНК с последовательностью нуклеотидов, соответствующей тому или иному гену (см.: РНК-интерференция).
В геноме рисовой цикадки имеются гены двух инсулиновых рецепторов (InR1, InR2) и четырех инсулиноподобных пептидов (ilp1, ilp2, ilp3, ilp4). Авторы начали с рецепторов. Оказалось, что отключение InR1 заставляет большинство подопытных личинок превращаться в короткокрылые взрослые формы, тогда как отключение InR2 дает противоположный эффект: почти все насекомые становятся в этом случае длиннокрылыми.
Из четырех инсулиноподобных пептидов только один (ilp3) оказался непосредственно задействован в регуляции полифенизма. Отключение гена, кодирующего этот пептид, повышает вероятность развития короткокрылых форм. Отключение остальных трех генов не влияет на длину крыльев. Авторы показали, что ilp3 синтезируется в мозге.
Дополнительные эксперименты показали, что инсулиноподобный пептид ilp3 взаимодействует с рецептором InR1, который в ответ на этот сигнал фосфорилирует протеинкиназу Akt, переводя ее в активное состояние. Активированная протеинкиназа, в свою очередь, фосфорилирует транскрипционный фактор FOXO. Этот белок, будучи фосфорилированным, теряет активность и перестает транспортироваться в ядро. Попадая в ядро, активированный FOXO включает какие-то (пока не установленные) гены, не позволяющие крыльям нормально развиться (или, может быть, наоборот — выключает гены, необходимые для развития крыльев). Таким образом, «по умолчанию» цикадка развивается по длиннокрылому пути: инсулиноподобный пептид ilp3 взаимодействует с рецептором InR1, рецептор активирует киназу Akt, киназа инактивирует транскрипционный фактор FOXO, транскрипционный фактор не попадает в ядро, и крылья развиваются нормально.
Однако всё меняется, если в процесс вмешивается второй инсулиновый рецептор, InR2. Он очень похож по своей аминокислотной последовательности на InR1, но выполняет прямо противоположную функцию. Второй рецептор взаимодействует с первым и выводит его из строя, не позволяя активировать киназу Akt в ответ на инсулиновый сигнал. Таким образом, если в зачатках крыльев у нимфы активен белок InR2, развитие идет по короткокрылому пути: InR1 не активирует киназу, киназа не выводит из строя транскрипционный фактор FOXO, последний проникает в ядро и блокирует развитие крыльев (рис. 2).
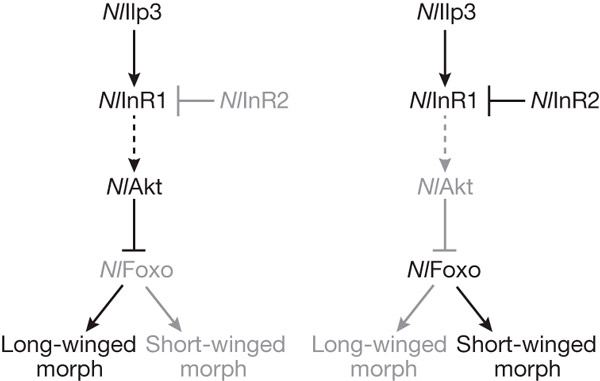
Рис. 2. Схема сигнального каскада, регулирующего развитие крыльев у рисовой цикадки. Стрелочками показаны положительные воздействия (активация), тупичками — отрицательные (ингибирование, инактивация). Буквы Nl перед названиями белков соответствуют латинскому названию рисовой цикадки, Nilaparvata lugens. Слева: рецептор inR2 неактивен, развитие идет по «длиннокрылому» пути (Long-winged morph). Справа: inR2 активен, развиваются короткокрылые формы (Short-winged morph). Рисунок из обсуждаемой статьи в Nature
По-видимому, внешние факторы каким-то образом влияют на активность InR2 в зачатках крыльев. Именно этим определяется судьба зачатков: станут ли они полноценными крыльями или так и останутся никуда не годными зачатками. Конкретный механизм влияния внешних факторов на активность InR2 пока неизвестен: будущие исследования должны прояснить этот вопрос.
Гомологи всех упомянутых генов и белков (инсулиноподобных пептидов, инсулиновых рецепторов, Akt и FOXO) есть и у человека, и у дрозофилы (см., например: FOXO1). Это классические участники инсулиновых сигнальных каскадов, регулирующих метаболизм глюкозы, клеточный цикл, рост органов и тканей и многие другие процессы, протекание которых варьирует в зависимости от количества питательных веществ в организме. По-видимому, у цикадок этот сигнально-регуляторный каскад просто взял на себя еще одну дополнительную функцию. С эволюционно консервативными, многофункциональными генами-регуляторами такое происходит сплошь и рядом. О других примерах появления новых функций у древних регуляторных генов мы рассказывали в новостях: Найден ген, отвечающий за эволюцию окраски у бабочек, «Элементы», 31.08.2011; Найден ген, превращающий простые листья в сложные, «Элементы», 18.02.2014.
Источник: Hai-Jun Xu, Jian Xue, Bo Lu, Xue-Chao Zhang, Ji-Chong Zhuo, Shu-Fang He, Xiao-Fang Ma, Ya-Qin Jiang, Hai-Wei Fan, Ji-Yu Xu, Yu-Xuan Ye, Peng-Lu Pan, Qiao Li, Yan-Yuan Bao, H. Frederik Nijhout & Chuan-Xi Zhang. Two insulin receptors determine alternative wing morphs in planthoppers // Nature. 2015. V. 519. P. 464–467.
См. также о полифенизме:
1) Выведена гусеница, меняющая цвет при нагревании, «Элементы», 09.02.2006.
2) Параллельная эволюция касты «сверхсолдат» у муравьев получила объяснение, «Элементы», 15.01.2012.
3) Самцы и самки меняются ролями при смене погоды, «Элементы», 27.12.2012.
-
Крыловой полифенизм - очень неудачная калька. Полифенизм - явление многогранное, и при желании можно найти различия в любых тканях, органах и функциях особей разных фенов (фаз, морф, морфотипов и т.п.) По-русски стоит писать только полифенизм крыльев, т.к. на них заострено внимание в данной статье (мышцы, половые железы, гиподерма, мозги, поведение - по боку).
< Удалено/отредактировано >
Название вводит в заблуждение почти всех читателей: "новое" не относится ни к функции (давным давно описана роль I/IGF в морфогенезе), ни к объекту - примерно так всё устроено у Pancrustacea. На других организмах либо нет данных, либо нет сравнимой морфологии (нематоды), но эндокринология Ecdysozoa вообще очень консервативна.
А статья-то добротная и кропотливая, но вообще слабенькая по своей сути: годичная выдержка говорит, что Nijhout был привлечён для коррекции текста и облагораживания коллектива... И наверняка что-то переделывать заставили.
Механизм прикольный.


Последние новости








Рис. 1. Короткокрылая и длиннокрылая формы бурой рисовой цикадки Nilaparvata lugens. Изображение из обсуждаемой статьи в Nature