Общий предок вторичноротых мог быть похож на хордовое

Проблема происхождения уникальной трубчатой центральной нервной системы хордовых животных до сих пор не решена. Американский биолог Линда Холланд сделала обзор современного состояния этой темы, привлекая новые данные из области генетики развития. В итоге она склоняется к гипотезе происхождения нервной трубки хордовых от брюшного нервного тяжа древних двусторонне-симметричных животных и делает вывод, что общий предок вторичноротых, вероятно, был очень похож на хордовое.
Один из главных признаков типа хордовых состоит в том, что нервная система у них (вернее, у нас) устроена в виде трубки. Очень часто центральную нервную систему хордовых животных так и называют: нервная трубка (рис. 1). Этот признак — уникальный. Ни у каких других животных трубчатой центральной нервной системы нет. Откуда же она у хордовых взялась?
Удивительно, но на этот вопрос до сих пор нет никакого общепринятого ответа. И это при том, что нервная трубка входит в характеристику хордовых, которая есть в каждом учебнике зоологии. Происхождение центральной нервной системы хордовых — научная загадка, сохраняющая актуальность почти двести лет, со времен великого француза Этьена Жоффруа Сент-Илера (Étienne Geoffroy Saint-Hilaire). Во времена, когда Сент-Илер над этим задумывался, еще и названия «хордовые» не существовало, а было только «позвоночные».
Американский биолог Линда Холланд (Linda Holland), сотрудница Института океанографии имени Эллен Скриппс (Scripps Institution of Oceanography) при Калифорнийском университете в Сан-Диего и крупный специалист по ранней эволюции хордовых, опубликовала обзор современного состояния проблемы происхождения их ЦНС, дополнив его собственными новыми идеями.
Положение хордовых на эволюционном древе животных, по современным данным, таково. Крупная ветвь, к которой они принадлежат, называется вторичноротые (Deuterostomia). Кроме хордовых, к вторичноротым бесспорно относятся еще два типа: иглокожие и полухордовые. Они и есть самые близкие современные родственники хордовых. Такие животные, как членистоногие, моллюски и черви, никакого прямого отношения к хордовым не имеют — они образуют ветвь первичноротых (Protostomia), гораздо более далекую от нас.
Сами вторичноротые распадаются на две главные эволюционные ветви (рис. 2). Одна ветвь называется Ambulacraria, в нее входят иглокожие и полухордовые. Важный общий признак этих животных — планктонная личинка, плавающая с помощью ресничек, которые собраны в так называемые ресничные шнуры. Ранние личинки иглокожих и полухордовых настолько похожи, что для них придумано общее название: диплеврула. Вторая главная ветвь вторичноротых — это и есть хордовые (Chordata). Ни у кого из них личинки-диплеврулы нет.

Рис. 2. Современное эволюционное древо вторичноротых. Пояснения в тексте
Хордовые, в свою очередь, делятся еще на три ветви. К одной из них принадлежим мы с вами — это позвоночные. Вторая ветвь называется бесчерепные, из современных животных к ней относится только ланцетник. И третья — оболочники, они же личиночнохордовые, существа очень необычного облика, у которых хорду чаще всего имеет только личинка (см. Причина особенностей генома оболочников — детерминированность их эмбрионального развития, «Элементы», 01.06.2014).
По современным данным, ближайшими родственниками позвоночных являются оболочники. Группа, которую они вместе образуют, называется Olfactores (букв. «животные с органом обоняния»). Ланцетник в эту группу не входит, его эволюционная веточка отошла от общего ствола хордовых раньше.
Что касается версий происхождения центральной нервной системы хордовых, то они довольно разнообразны; иначе и не могло быть, учитывая, насколько эта проблема старая. В целом, идеи, существующие на эту тему, можно свести к пяти главным гипотезам.
Гипотеза происхождения от ресничных шнуров. Исходит из того, что общий предок всех вторичноротых был во взрослом состоянии похож на личинку-диплеврулу (рис. 3, А). Пара ресничных шнуров этого «диплеврулообразного» предка срослась на средней линии, образовав сначала нервную пластинку, а потом и нервную трубку. Действительно, наша нервная трубка до сих пор выстлана изнутри слоем ресничных клеток (эпендимой). Однако личинка-диплеврула есть не у всех вторичноротых, а только у представителей ветви Ambulacraria. У Chordata диплеврулы нет, и нет никаких свидетельств, что она у них когда-то была. В такой ситуации мнение, что предок хордовых проходил «диплеврулообразную» эволюционную стадию, выглядит слабо обоснованным. Не менее вероятно, что диплеврула — это уникальная особенность Ambulacraria, то есть иглокожих и полухордовых. Тогда к происхождению хордовых ее устройство не имеет никакого отношения.

Рис. 3. Гипотезы происхождения хордовых от диплеврулообразного предка (А) и от радиально-симметричного предка (Б). Зеленым показаны нервные структуры и ресничные шнуры, от которых они предположительно произошли. Эндостиль — желобок на дне глотки, выстланный ресничными клетками. Остальные пояснения в тексте. Рисунки из обсуждаемой статьи в The Journal of Experimental Biology, с изменениями
Гипотеза происхождения от брюшного нервного тяжа. Очень старая идея (впервые высказана еще Сент-Илером), согласно которой нервная трубка хордовых соответствует брюшной нервной цепочке первичноротых, то есть членистоногих и кольчатых червей. Нервная трубка расположена на спине, а брюшная нервная цепочка — естественно, на брюхе. Следовательно, в какой-то эволюционный момент предок хордовых в буквальном смысле перевернулся брюхом кверху. Предположение, что спинная сторона хордовых соответствует брюшной стороне всех остальных животных, удивительно хорошо подтверждается современной молекулярной биологией развития (см., например: Д. А. Воронов, 2000. Старая гипотеза «перевернутости» хордовых подтверждается). Чаще всего считается, что центральная нервная система возникла раньше, чем спинная и брюшная стороны тела поменялись местами; хотя, возможно, брюшная нервная цепочка тогда была еще не цепочкой (состоящей из ганглиев), а просто тяжом.
Гипотеза независимой концентрации. Альтернатива предыдущей гипотезе, предполагающая, что у общего предка хордовых и первичноротых нервная система была диффузной, то есть охватывала тело равномерной сетью. Превращение диффузной нервной системы в центральную — часть эволюционного процесса, который обычно называют концентрацией. Возможно, концентрация нервной системы произошла независимо у хордовых (на стороне, которая стала спинной) и у ряда других животных (на стороне, которая стала брюшной). Главная особенность этой гипотезы — она предполагает гораздо более простое устройство организма предка хордовых, чем гипотеза происхождения нервной трубки от брюшного нервного тяжа.
Гипотеза радиально-симметричного предка. Довольно редкая идея, согласно которой предок хордовых имел радиальную (лучевую) симметрию — примерно как иглокожие (хотя и не обязательно сам был иглокожим). Например, такая симметрия могла быть пятилучевой (рис. 3, Б). В таком случае предок хордовых должен был иметь пять нервных тяжей — по одному в каждом луче. Когда симметрия сменилась на двустороннюю, четыре нервных тяжа исчезли, а единственный оставшийся разросся и стал нервной трубкой. Совсем недавно с такой гипотезой выступил известный биолог-эволюционист, сотрудник МГУ Александр Николаевич Кузнецов (A. N. Kuznetsov, 2012. Five longitudes in chordate body). Работа Кузнецова — спорная, но интересная — была поначалу обойдена вниманием западных ученых. Линда Холланд в обсуждаемой статье цитирует ее чуть ли не впервые. Увы, гипотеза радиально-симметричного предка хордовых с трудом согласуется с данными молекулярной биологии развития, и это, видимо, сейчас основная причина ее непопулярности.
Гипотеза нервной трубки у полухордовых. Еще в XIX веке было обнаружено, что трубчатый участок, очень похожий на нервную трубку хордовых, есть в нервной системе морских червеобразных животных, относящихся к типу полухордовых (рис. 4). Правда, «нервная трубка» полухордовых гораздо короче, она тянется далеко не на все тело. Тем не менее, было очень соблазнительно предположить, что хордовые произошли от полухордовых и что трубчатое устройство, имеющееся у полухордовых в зачаточном состоянии, потом распространилось на всю центральную нервную систему. Некоторые ученые поддерживают такую идею и сейчас. Но с этим связаны две проблемы. Во-первых, полухордовые — не предки хордовых. По современным представлениям, степень родства с хордовыми у полухордовых и у иглокожих строго одинакова, поэтому трудно сказать, на кого их общий предок был больше похож. Во-вторых, трубчатый участок нервной системы полухордовых расположен на той стороне тела, которая соответствует брюху хордовых, а вовсе не спине; по крайней мере, данные молекулярной биологии развития однозначно на это указывают. Поэтому сейчас более распространено мнение, что трубчатые нервные структуры появились у хордовых и у полухордовых независимо.

Рис. 4. Строение полухордового Saccoglossus kowalevskii. Иллюстрация из статьи: C. Lowe et al., 2003. Anteroposterior Patterning in Hemichordates and the Origins of the Chordate Nervous System (с изменениями)
Подводя итог, можно сказать, что актуальнее всего теперь две гипотезы: гипотеза происхождения нервной трубки от брюшного нервного тяжа и гипотеза независимой концентрации. Окончательный выбор между ними еще не сделан.
Полухордовые упомянуты здесь не случайно. Для всех проблем, связанных с происхождением хордовых, они в любом случае представляют совершенно исключительный интерес. Это единственные современные родственники хордовых, с которыми последних можно сравнить. Конечно, есть еще иглокожие, но у тех план строения видоизменен буквально фантастически, так что от сравнений мало толку.
Увы, с нервной системой у полухордовых все неоднозначно. С одной стороны, у них есть подкожная нервная сеть, охватывающая все тело. Такую нервную систему вполне можно считать диффузной. С другой стороны, нервная сеть полухордовых кое-где сгущается, образуя тяжи. Причем продольных нервных тяжей у них два: на спине и на брюхе. Какой из них соответствует нервной трубке хордовых и имеет ли вообще смысл такой вопрос, до сих пор не совсем ясно. Данные по полухордовым можно истолковать как в пользу гипотезы происхождения нервной трубки от брюшного нервного тяжа, так и в пользу гипотезы независимой концентрации; и действительно, есть ученые, трактующие их и так и эдак.
Линда Холланд обращает внимание еще и на то, что нервная система полухордовых обладает возрастной изменчивостью. Для их самого изученного представителя — Saccoglossus kowalevskii — есть данные, что у зародышей до вылупления нервная система ближе к диффузной, а затем она частично «централизуется» (D. Cunningham, E. Casey, 2014. Spatiotemporal development of the embryonic nervous system of Saccoglossus kowalevskii). Говорит ли это что-нибудь об эволюционном пути полухордовых, пока неизвестно.
И наконец, Линда Холланд по примеру уже многих исследователей привлекает к вопросу об эволюции нервной системы данные генетики развития. Есть гены, экспрессия (проще говоря, активность) которых привязана или к спинной, или к брюшной стороне тела. Например, у зародыша ланцетника ген BMP2/4 (BMP означает bone morphogenetic protein, «морфогенетический белок костей») экспрессируется на брюшной стороне и сзади, а эволюционно относительно близкий к нему ген Nodal — на спинной стороне и впереди. Продукты обоих генов — сигнальные белки, действующие на разные (хотя и похожие) клеточные рецепторы. Физиологически гены BMP2/4 и Nodal являются антагонистами, то есть подавляют действие друг друга. Функция гена Nodal связана с развитием центральной нервной системы, расположенной у хордовых на спине. Избыточная активность этого гена приводит к непомерному расширению зачатка нервной системы и превращению почти всего внешнего слоя клеток зародыша в подобие нервной пластинки (skin-brain, «кожа-мозг»). Избыточная активность гена BMP2/4, наоборот, ведет к полному отсутствию центральной нервной системы, а заодно и головы. Генов BMP у животных обычно несколько, отсюда и цифры в названии указанного гена у ланцетника.
Есть основания думать, что такое разделение функций между генами BMP и Nodal существовало уже у общего предка всех двусторонне-симметричных животных, как первичноротых, так и вторичноротых. Правда, на любимом объекте генетиков — мухе-дрозофиле — проверить это нельзя, так как гена Nodal у нее просто нет. Но у кольчатых червей есть и ген BMP, и ген Nodal, и похоже, что они взаимодействуют примерно так же, как у хордовых. Кольчатые черви, безусловно, относятся к первичноротым. А это, при данной конфигурации эволюционного древа, означает, что общий предок кольчатых червей и хордовых мог быть только общим предком всех двусторонне-симметричных животных (см., например: Новые данные позволили уточнить родословную животного царства, «Элементы», 10.04.2008). Если система BMP-Nodal унаследована от него — значит, она очень древняя.
Но вот у полухордовых эта система работает иначе. Правда, экспрессия BMP у них традиционно привязана к определенной стороне тела, которая считается спинной (Saccoglossus kowalevskii, на котором это показано, живет в норах и проводит большую часть своей жизни в вертикальном положении, так что само выделение спинной и брюшной сторон тут довольно условно). Однако экспрессия Nodal у полухордовых почему-то маркирует не «брюшную» сторону (как можно было бы ожидать), а заднюю часть тела, без особой связи со спиной или брюхом. Линда Холланд предполагает, что в эволюционной линии полухордовых взаимодействие между генами BMP и Nodal, основанное на их антагонизме, оказалось по какой-то причине потеряно. Эта потеря увеличила число степеней свободы в организме полухордовых, позволив там возникнуть дополнительному нервному тяжу, который появился на так называемой «спинной» стороне тела — в зоне экспрессии BMP, где у большинства животных нет никаких нервных структур.
Линда Холланд, как видим, поддерживает гипотезу происхождения нервной трубки от брюшного нервного тяжа. Она подчеркивает, что в рамках предлагаемого сценария потеря антагонизма «BMP versus Nodal» должна быть вторичной (рис. 5). У полухордовых «сломался» механизм, который у их предков был. Как же, в таком случае, выглядели эти предки?
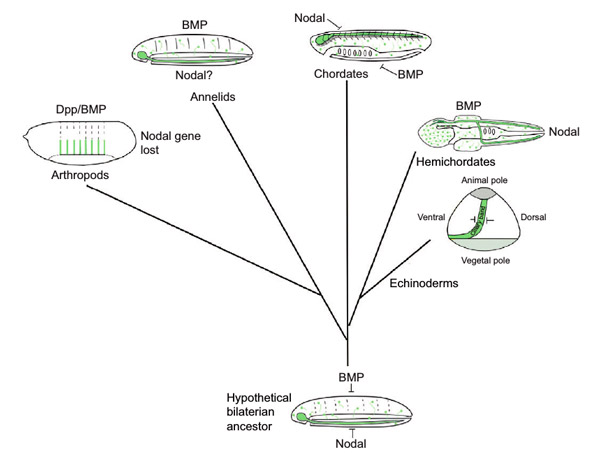
Рис. 5. Сценарий эволюции нервной системы, предложенный Линдой Холланд. У гипотетического предка двусторонне-симметричных животных (Hypothetical bilaterian ancestor) на брюшной стороне экспрессировался ген Nodal, на спинной ген BMP, и в области экспрессии Nodal развивался брюшной нервный тяж. Гены Nodal и BMP были антагонистами, то есть подавляли действие друг друга. У кольчатых червей (Annelids) эта система сохраняется в том же виде. У членистоногих (Arthropods) она перестраивается из-за потери гена Nodal; ген BMP (он же Dpp) у них сохраняет прежнюю функцию. У иглокожих (Echinoderms) антагонистическая экспрессия Nodal и BMP предположительно регулирует закладку личиночных ресничных шнуров (Ciliary band); анимальный полюс (Animal pole) и вегетативный полюс (Vegetative pole) — физиологические полюса раннего зародыша. У полухордовых (Hemichordates) экспрессия Nodal смещается назад, антагонизм между Nodal и BMP исчезает. Возможно, именно это нарушение взаимодействия каким-то образом приводит к появлению дополнительного — спинного — нервного тяжа в области экспрессии BMP. И, наконец, хордовые наследуют систему взаимодействия Nodal и BMP прямо от общего предка двусторонне-симметричных животных, в мало измененном виде, только вот спинная и брюшная стороны у них меняются местами. Иллюстрация из обсуждаемой статьи в The Journal of Experimental Biology
У всех животных, у которых отмечен (или предполагается) антагонизм «BMP versus Nodal», есть центральная нервная система, расположенная на той стороне, где экспрессируется Nodal. Вероятно, это связанные признаки. Тогда получается, что у общего предка вторичноротых, скорее всего, была центральная нервная система. А это означает, что общий предок вторичноротых был больше похож на хордовое, чем на представителя любого другого известного типа.
Нельзя не заметить, что эта версия отлично согласуется с некоторыми другими новыми данными: например, с открытием органа, напоминающего хорду, у кольчатых червей (см. У предков билатерий уже была примитивная протохорда, «Элементы», 18.09.2014). Возможно, что общий предок не только вторичноротых, но и всех двусторонне-симметричных животных был очень похож на хордовое. Эта гипотеза еще лет двадцать назад выглядела бы поразительно, но сейчас она рассматривается всерьез.
Линда Холланд приводит еще один факт, который нельзя считать серьезным свидетельством, но можно воспринять как повод задуматься. В прошлом году вышла нашумевшая статья международной группы авторов, главной целью которых было выяснить происхождение нервной системы гребневиков (L. Moroz et al., 2014. The ctenophore genome and the evolutionary origins of neural systems). Обзор этой статьи тогда же появился на «Элементах» (Гипотеза о двукратном появлении нервной системы получила новые подтверждения, «Элементы», 26.05.2014). Работа была в основном генетической. Чтобы разобраться в «сюжете», интересовавшем авторов, им пришлось построить по своим данным эволюционные деревья, охватывающих всех животных в целом, в том числе и вторичноротых. Как обычно в таких работах, деревьев было построено несколько, с небольшим варьированием выборок данных и методических подходов. Так вот, среди полученных схем оказалась такая, в которой эволюционная ветвь ланцетника почему-то примыкает не к группе Olfactores, а к группе Ambulacraria (см. рис. 2). Если бы такое древо оказалось верным, оно означало бы, что хордовые животные есть в обеих крупных ветвях вторичноротых. А это уже точно вело бы к выводу, что общий предок вторичноротых был хордовым; сходство ланцетника и позвоночных слишком велико, чтобы предполагать независимое происхождение такого большого набора признаков.
Всерьез это воспринимать пока не стоит, ибо древо, где ланцетник ближе к полухордовым и иглокожим, чем к другим хордовым, совсем не общепринято. Но бывает, что и сомнительные факты наводят на продуктивные идеи.
При всем этом Линда Холланд сама оговаривает «спекулятивный» (по ее выражению) характер предлагаемого эволюционного сценария. Вопрос далеко не закрыт.
Ну, а почему же все-таки нервная система хордовых имеет форму трубки? Этого никто не знает. Возможно, все дело в том, что эта нервная система изначально была чрезвычайно близка к коже; тогда самым простым способом сделать из нее плотную структуру было сворачивание покровного пласта в трубочку, что мы и видим в спинном нервном тяже полухордовых. Даже если нервная трубка хордовых возникла независимо, причина ее «трубчатости» может быть той же самой. Но ответить на этот вопрос четко можно будет лишь тогда, когда проблема происхождения ЦНС хордовых будет решена в принципе — то есть будет сделан окончательный выбор из перечисленных в нашем обзоре пяти гипотез. На данный момент это еще не случилось.
Источник: Linda Z. Holland. Evolution of basal deuterostome nervous systems // The Journal of experimental biology.2015. V. 218. № 4. P. 637–645.
Сергей Ястребов
-
>Возможно, что общий предок не только вторичноротых, но и всех двусторонне-симметричных животных был очень похож на хордовое. Эта гипотеза еще лет двадцать назад выглядела бы поразительно, но сейчас она рассматривается всерьез.
Похоже, мы становимся свидетелями (а некоторые - и участниками) определенного изменения парадигмы филогенетики: что если мы, приученные к осторожности в выводах бурностью развития биологии последнего полувека, проявляем её порой и на ровном месте, всегда говоря о "гипотетическом общем предке", тогда как он - вполне вероятно - практически в неизменном виде плавает вокруг нас или сохраняется внутри нас??
И у Е.Кунина в книге проходит аналогичная идея в отношении происхождения и митохондрий и жизни вообще: может быть пелагибактер действительно за прошедшие два миллиарда лет почти не изменился с момента когда он стал предком митохондрий, а рибосома действительно содержала большую часть всего необходимого для репликации (http://elementy.ru/news/432415). Так и ланцетник (тоже весьма убиквитарное существо, кстати!) - с кембрия ушел совсем-совсем недалеко от общего предка вторичноротых?
Этот подход становится всё более убедительным в свете ещё и таких данных: http://postnauka.ru/faq/42341
Может быть через десяток-другой лет фразы для широкого круга неспециалистов типа "митохондрии произошли от пелагибактера", "рибосома - самый древний и центральный органоид первых клеток", "все вторичноротые произошли от зверушки, очень похожей на ланцетника (в разрезе)" не будут вызывать бурю негодования у специалистов даже если они окажутся в школьных учебниках биологии?-
ну тогда если уж пошла такая пляска сбросим оковы науч.корректности и скажем открыто "человек произошел от обезьяны"
-
-
по-разному можно сказать кто от кого и как именно произошел
но обычно считается что указание на прямое линейное происхождение одного от другого ненаучно без железобетонных доказательств
соответственно используется более мягкая формулировка "имели (близкого) общего предка"
"человек произошел от обезьяны" - редукция научного тезиса и запутанного биологического явления до легко усваиваемого медийного мема
аналогично предлагаемое упрощение о "происхождении митохондрий от пелагибактера" в научном смысле (для специалистов) не добавляет ничего нового но упрощает учебники превращая в пособия для кухарок-
-
-
-
-
-
когда Дарвин опубликовал свой труд недалёкая публика в течение десятилетий мусолила вопрос о каких таких обезьянах говорят эти дарвинисты. и не без успеха мусолила. Потому что среди многих чванливых ученых не укоренилась мысль, что с термилогией - monkeys, apes, the apes, extinct apes, living apes и т.д. - надо быть поаккуратнее, особенно с людьми, жадно ловящих каждый прокол эволюционистов.
Религиозная и просто малообразованная публика вполне обоснованно в течение столетий уже рисует карикатуры на произошедших от макак, а вы всё во главе с И.Ивановым боретесь здесь с "альтернативщиками".
Это конечно абсолютно мелкий и частный вопрос, и удивительно, что всем в этой ветке что-то оказалось непонятным в моем самом первом замечании, и все решили последовательно выводить идиота на чистую воду. Поздравляю, блестяще справились, ребята!-
1. По-русски и monkey, и ape - обезьяна. Из всех родословных древес, кроме совсем уже неправдоподобных, следует, что общий предок человека и шимпанзе был обезьяной (скорее всего, даже человекообразной). Никакой неоднозначности, никакой зависимости от варьируемых значений терминов тут нет. Человек произошел от обезьяны. Я об этом уже мимоходом писал вот тут:
http://elementy.ru/news/432416
2. Я не знаю, кто такой И. Иванов и кто такие "альтернативщики".
3. Мне жаль, если вы приняли что-то в дискуссии на свой счет. Ничего подобного не предполагалось, я честно задавал уточняющие вопросы, как задал бы их любому собеседнику.
-
-
-
-
-
-
-
-
-
-
Учитывая систематику человек как был обезьяной так ею и остался
Отряд: Приматы
Подотряд: Сухоносые обезьяны
Инфраотряд: Обезьянообразные
Парвотряд: Узконосые обезьяны
Надсемейство: Человекообразные обезьяны
Семейство: Гоминиды
Подсемейство: Гоминины
Триба: Хоминини
Подтриба: Хоминина
Род: Люди
Вид: Человек разумный
-


"Этот признак — уникальный. Ни у каких других животных трубчатой центральной нервной системы нет." Это странным образом противоречит дальнейшему содержанию заметки. Ведь нервная трубка полухордовых - часть их ЦНС, разве не так?
"Вторая ветвь называется бесчерепные, из современных животных к ней относится только ланцетник." Может быть, лучше было бы ланцетники? Это как-никак несколько родов с довольно большим числом видов и не такой уж однообразной морфологией...
"У Chordata диплеврулы нет, и нет никаких свидетельств, что она у них когда-то была." Вообще-то у ланцетника из яйца выходят ресничная личинка, имеющая три пары целомов (задние к этому моменту разделены на пару сомитов) и сквозной кишечник. От диплеврулы она отличается в основном строением нервной системы (ну, и еще тем, что ресничный покров у нее сплошной). По-моему, это прямое свидетельство того, что у предков хордовых была диплеврулоподобная личинка.
"Во-вторых, трубчатый участок нервной системы полухордовых расположен на той стороне тела, которая соответствует брюху хордовых, а вовсе не спине; по крайней мере, данные молекулярной биологии развития однозначно на это указывают." Как следует даже из дальнейшего текста заметки, может быть, не так уж однозначно? Если взаимодействие между генами BMP и Nodal нарушено и область экспресси Nodal могла сместиться, то, возможно, могла сместиться и область экспрессии BMP (хотя, конечно, мне тоже кажется более вероятным, что спинная сторона тела кишечнодышащих соответствует брюшной стороне тела ланцетника).
"Но у кольчатых червей есть и ген BMP, и ген Nodal, и похоже, что они взаимодействуют примерно так же, как у хордовых." Если это данные из статьи - то там написано еще более уклончиво, и не случайно на рис.5 стоит знак вопроса рядом с Nodal у полихеты. Или есть еще какие-то дополнительные данные? Кстати, на рис. 5 и в заметке, и в статье выпали обозначения генов BMP и Nodal у иглокожих, хотя в тексте статьи их кспрессия описывается...
Жаль, в заметке уделено мало внимания развитию нервной итемы ланцетника. Например, написано, что "Избыточная активность гена BMP2/4, наоборот, ведет к полному отсутствию центральной нервной системы, а заодно и головы." Но не написано, что при той же избыточной активности (http://dev.biologists.org/content/139/11/2020.full) во всей эктодерме закладываются периферические сенсорные нейроны, которые у ланцетника в норме закладываются на брюшной стороне тела...А можно было бы порассуждать, какую из гипотез этот факт скорее поддерживает.
И, конечно, еще один интересный вопрос - это строение и развитие ЦНС ксентурбеллы. Все-таки это, скорее всего, тоже амбулакрария - и неплохо было бы выяснить, как дело обстоит у нее. По-моему, ее стоило упомянуть, даже несмотря на отсутствие данных по развитю ее ЦНС к настоящему моменту.
-
1. "Это странным образом противоречит дальнейшему содержанию заметки. Ведь нервная трубка полухордовых - часть их ЦНС, разве не так?"
Нет, не так. У полухордовых _нет_ ЦНС. Если только не называть вообще любую нервную систему центральной, но тогда это понятие теряет смысл.
2. "Может быть, лучше было бы ланцетники?"
Может быть. Формально вы, конечно, правы. Хотя, с другой стороны, в популярной статье, где разнообразие бесчерепных никак не затрагивается, может быть, как раз и стоило выразиться грубее, но проще и понятнее. Еще подумаю над этим.
3. О диплевруле. Если у нее нервная трубка - то какая же это диплеврула? Если ресничный покров, как и вы подтверждаете, сплошной, без всяких ресничных шнуров - то какая же это диплеврула? И если случилось разделение заднего целома "на пару сомитов", то это уже не трехцеломная стадия. Нельзя же любую ресничную личинку называть диплеврулой! Я прекрасно понимаю, откуда эта трактовка взялась: в начале 20 века была популярно представление о диплевруле как об архетипе вторичноротых, о чем я и пишу, собственно. Но сейчас оснований так думать гораздо меньше.
Если хотите меня переубедить, это очень легко. Найдите любую статью, желательно опубликованную в последние 25 лет, с описанием и изображением личинки ланцетника, где было бы показано ее сходство с диплеврулой. Мне самому интересно. Тема популярная, пропустить такое исследователи не могли.
3. По экспрессии генов дорсо-вентральной полярности у полухордовых есть еще и другие данные, здесь не обсуждаемые:
http://caenogenesis.livejournal.com/50062.html
http://caenogenesis.livejournal.com/50237.html
Гомология сторон тела (не нервных тяжей, а именно сторон тела) тут, как мне кажется, подтверждена достаточно надежно.
4. Про Nodal у кольчатых червей дополнительных данных я не нашел, видимо, эту тему только начали исследовать. Здесь скорее предположение, сделанное Линдой Холланд, но мне оно кажется обоснованным.
5. Иглокожих я здесь сознательно стараюсь обсуждать по минимуму, уж слишком много у них уникальных признаков. И сам запутаюсь, и читателя запутаю. Про них лучше отдельно.
6. Про влияние гиперэкспрессии BMP у ланцетника на закладку сенсорных нейронов действительно очень интересно, это может иметь что-то общее с эволюцией полухордовых. Но столько тем - не для одной статьи. Она и так перегружена, обратите внимание на объем.
7. Про ксенотурбеллу я в ближайшее время (скорее всего, уже завтра) начну писать в научном блоге, на который даны ссылки выше. Из этой статьи я сознательно исключил разговор о ней, т.к. она (статья), на мой взгляд, и так на грани того, что может сходу воспринять свежий читатель. Нельзя же объять необъятное. Вот попадется хорошая новость про ксенотурбеллу, тут-то я ее и освещу... а пока обойдемся блогом.-
1. По первому пункту - может быть, тут Вас тоже убедят статьи?
Nomaksteinsky M., Röttinger E., Dufour H., Chettouh Z., Lowe C. J., Martindale M., Brunet J. 2009. Centralization of the deuterostome nervous system predates chordates. Curr. Biol. 19, 1264–1269
Luttrell S., Bengtsson B. C., Swalla B. J. 2010. Central nervous system development and regeneration in hemichordates. Integr. Comp. Biol. 50(Suppl. 1), e105
В первой статье логично обосновывается, почему у полухордовых есть ЦНС. Вторую статью я не нашел, но название говорит само за себя...
Встречный вопрос: если нервные тяжи и нервная трубка кишечнодышащих и крыложаберных - это не ЦНС, то что же такое тогда ЦНС?
3. Я же не говорю, что у ланцетника есть типичная диплеврула. Я возражаю против того, что у хордовых нет признаков "диплеврулообразных" личинок. Если у ланцетника из яйца выходит трехсегментная ресничная личинка - это указание на то, что у его предков могла быть личинка диплеврула, разве не так? А сегметация задних целомов у ланцетника начинается раньше, чем обособляются передние две пары целомов - ну и что с того?
Что касается статей - есть статьи Ивановой-Казас 1990-х годов, где она выводит хордовых из предков с диплеврулоподобной личинкой. Это , конечно, не статьи с рисунками, но возникшие тоже не на пустом месте. Зато из статей "с рисунками" нашлась статья Незлина (Tornaria of hemichordates and other dipleurula-type larvae: a comparison), где он пытается доказать, что сходство торнарий с личинками иглокожих конвергентное, а на самом деле их нервные системы устроены принципиально по-разному. Если это так - что же тогда считать диплеврулой?
В целом - спасибо за подробный ответ. За постами про ксенотурбеллу буду следить.-
Мне очень интересно обсудить с вами эту тему, но я вынужден взять тайм-аут, потому что уезжаю на десять дней туда, где связь будет так себе. Пока что кратко.
1. Может быть, и убедят, посмотрим. Пока я опираюсь вот на что:
http://caenogenesis.livejournal.com/46508.html
http://caenogenesis.livejournal.com/48567.html
http://caenogenesis.livejournal.com/50511.html
http://caenogenesis.livejournal.com/50924.html
http://caenogenesis.livejournal.com/50949.html
http://caenogenesis.livejournal.com/51361.html
А ЦНС, по-моему, - это скопление тел нейронов, преимущественно вставочных, морфологически выраженное в виде единственной компактной структуры.
2. Мне всегда казалось, что диплеврула, так же, как, например, трохофора, должна иметь определенные ресничные шнуры, гомологичные ресничным шнурам всех остальных диплеврул. Разве нет? Кроме того, я нигде не видел четкого доказательства, что у ланцетника есть именно трехсегментная личинка.
3. Про Иванову-Казас давайте отдельно. Она была очень мудра, но все-таки в данном случае это интерпретации, а не факты.
На этом пропадаю на время. Вернусь - будет и ксенотурбелла, и все остальное.
-
-
Эволюция хордовых
-
 16.09.2024Открыто новое кембрийское позвоночноеСергей Ястребов • Новости науки
16.09.2024Открыто новое кембрийское позвоночноеСергей Ястребов • Новости науки -
 26.12.2016Эмбриональное развитие брахиопод проливает свет на природу первично- и вторичноротостиАлександр Марков • Новости науки
26.12.2016Эмбриональное развитие брахиопод проливает свет на природу первично- и вторичноротостиАлександр Марков • Новости науки
-
 02.06.2016Обнаружено фундаментальное сходство между развитием актинии и развитием позвоночныхЮлия Краус, Александр Марков • Новости науки
02.06.2016Обнаружено фундаментальное сходство между развитием актинии и развитием позвоночныхЮлия Краус, Александр Марков • Новости науки
-
 24.11.2015Геномы полухордовых проливают свет на эволюцию вторичноротыхАлександр Марков • Новости науки
24.11.2015Геномы полухордовых проливают свет на эволюцию вторичноротыхАлександр Марков • Новости науки
-
 05.11.2015Гомологи хорды широко распространены у первичноротыхСергей Ястребов • Новости науки
05.11.2015Гомологи хорды широко распространены у первичноротыхСергей Ястребов • Новости науки
-
 18.03.2015Общий предок вторичноротых мог быть похож на хордовоеСергей Ястребов • Новости науки
18.03.2015Общий предок вторичноротых мог быть похож на хордовоеСергей Ястребов • Новости науки
-
 04.03.2015У ланцетника тоже есть хрящЕлена Наймарк • Новости науки
04.03.2015У ланцетника тоже есть хрящЕлена Наймарк • Новости науки
-
 04.02.2015«Четвертый зародышевый листок» позвоночных зародился у низших хордовыхСергей Ястребов • Новости науки
04.02.2015«Четвертый зародышевый листок» позвоночных зародился у низших хордовыхСергей Ястребов • Новости науки
-
 18.09.2014У предков билатерий уже была примитивная протохордаЕлена Наймарк • Новости науки
18.09.2014У предков билатерий уже была примитивная протохордаЕлена Наймарк • Новости науки
-
 25.10.2013У одноклеточных организмов есть ген, способный управлять развитием хордыСергей Ястребов • Новости науки
25.10.2013У одноклеточных организмов есть ген, способный управлять развитием хордыСергей Ястребов • Новости науки
Последние новости











Рис. 1. План строения типа хордовых. Жирным шрифтом выделены признаки, которые считаются уникальными и традиционно входят в общую характеристику типа. Иллюстрация с сайта bio.utexas.edu (с изменениями)