Генетическое разнообразие популяции связано обратной зависимостью с уровнем заботы о потомстве

Виды животных сильно отличаются друг от друга по уровню генетического разнообразия (полиморфизма), однако причины этих различий точно не установлены. Анализ транскриптомов 76 видов животных, относящихся к 31 семейству и восьми типам, позволил выявить ключевой фактор, коррелирующий с уровнем генетического полиморфизма. Им оказался уровень родительского вклада в потомство, который можно оценить по размеру особей на той стадии, когда они покидают родителей и переходят к самостоятельной жизни. Как выяснилось, низкий генетический полиморфизм характерен для видов, выпускающих в мир немногочисленное, но зато крупное и способное за себя постоять потомство, а высокий — для тех, кто бросает многочисленных мелких, незащищенных потомков на произвол судьбы. Данный результат заставляет пересмотреть некоторые устоявшиеся представления популяционной генетики и по-новому взглянуть на эволюционную роль заботы о потомстве.
Уровень генетического полиморфизма популяции (или вида в целом) считается в популяционной генетике важнейшим показателем, от которого зависят эволюционная пластичность вида, его приспособляемость к переменам среды и риск вымирания.
Виды животных сильно различаются по уровню генетического полиморфизма. Например, у гепардов разнообразие крайне низкое. Это объясняется недавним «бутылочным горлышком» — экстремальным снижением численности, в результате которого почти весь предковый полиморфизм был потерян. Поэтому все ныне живущие гепарды являются близкими родственниками, а генетически они почти идентичны друг другу. У ланцетника, наоборот, полиморфизм рекордно высок (см.: Геном ланцетника помог раскрыть секрет эволюционного успеха позвоночных, «Элементы», 23.06.2008). Это, предположительно, объясняется тем, что численность популяции ланцетников оставалась очень высокой в течение долгого времени.
Впрочем, одной лишь численностью популяции невозможно объяснить межвидовые различия по уровню полиморфизма. Выдающийся эволюционный генетик Ричард Левонтин (Richard Lewontin) еще 40 лет назад назвал объяснение этих различий центральной проблемой популяционной генетики (R. C. Lewontin, 1974. The Genetic Basis of Evolutionary Change). Однако добиться полной ясности в данном вопросе пока не удалось.
В теории проблема выглядит относительно простой. Согласно нейтральной теории молекулярной эволюции, в «идеальной» популяции (с абсолютно свободным, случайным скрещиванием, постоянной численностью, равным числом самцов и самок и т. д.) должен поддерживаться постоянный, равновесный уровень нейтрального генетического полиморфизма, зависящий только от двух переменных: темпа мутагенеза (частоты появления новых нейтральных мутаций) и эффективной численности популяции, Ne (см. также Effective population size). Последняя в идеале равна числу особей, участвующих в размножении, но в далекой от идеала реальности вычислять ее приходится сложными окольными путями — например, по косвенным признакам, указывающим на силу генетического дрейфа: чем ниже Ne, тем сильнее должен быть дрейф (см. краткое содержание главы 3 “Effective population size” учебника по популяционной генетике).
Для большинства видов измерить Ne затруднительно. Гораздо проще оценить «обычную» численность (N). Поскольку Ne, по-видимому, все-таки должна (со всеми оговорками) положительно коррелировать с N, логично предположить, что у массовых видов генетическое разнообразие должно быть в среднем выше, чем у малочисленных.
Эмпирические данные, однако, не дают этой гипотезе однозначного подтверждения. Похоже, различия по N позволяют объяснить лишь небольшую долю межвидовой вариабельности по уровню полиморфизма. Чем же объясняется всё остальное? Большинство специалистов предполагают совокупное влияние множества факторов, таких как темп мутагенеза (прямые данные по которому есть пока лишь для немногих видов), популяционная структура и ее динамика, отбор полезных мутаций (приводящий к «выметанию» нейтрального полиморфизма из окрестностей мутантного локуса). Но главным фактором обычно считают историческую динамику численности, в том числе наличие в прошлом периодов резкого сокращения численности (как в случае с гепардами) или длительное их отсутствие (как в случае с ланцетниками).
Впрочем, до сих пор попытки эмпирическим путем выяснить причины межвидовых различий по уровню полиморфизма имели фрагментарный характер: анализировались либо отдельные группы животных, либо небольшое число генов. Коллектив генетиков из Франции, Великобритании, Швейцарии и США попытался найти более общее решение «центральной проблемы популяционной генетики» при помощи современных методов секвенирования транскриптомов. Авторы получили и проанализировали транскриптомы 76 видов животных, относящихся к разным ветвям эволюционного дерева. Изученные виды представляют 31 семейство животных, принадлежащих к восьми типам: нематодам, членистоногим, моллюскам, немертинам, кольчатым червям, иглокожим, хордовым и книдариям.
Всего было исследовано 374 транскриптома, то есть в среднем изучено примерно по пять особей каждого вида и по 10 копий каждого гена (поскольку особи диплоидные). Этого достаточно, чтобы с приемлемой точностью оценить уровень полиморфизма белок-кодирующих последовательностей. В качестве меры нейтрального полиморфизма авторы использовали стандартный показатель — процент синонимичных различий между двумя случайно выбранными гомологичными последовательностями, πs. Был вычислен также процент несинонимичных (значимых) различий πn (см. Nucleotide diversity).
Оказалось, что уровень полиморфизма в изученной выборке варьирует в широких пределах. Рекордно низкое генетическое разнообразие обнаружилось у термита Reticulitermes grassei (πs = 0,1%), максимальное — у морского брюхоногого моллюска Bostrycapulus aculeatus (πs = 8,3%). Различие почти на два порядка!
Виды с высоким и низким полиморфизмом распределены по эволюционному дереву довольно хаотично (рис. 1). При этом родственные виды (относящиеся к одному и тому же семейству) в среднем более сходны друг с другом по уровню полиморфзима, чем представители разных семейств. Этот факт противоречит гипотезе о том, что главным фактором, влияющим на полиморфизм, являются случайные перипетии популяционной истории. Ведь нет оснований предполагать, что у видов, относящихся к одному семейству, должна быть сходная динамика численности. Правда, тут мог сказаться и подбор видов для анализа: например, все три вида морских ежей семейства Schizasteridae, выбранные для анализа, — это обитатели высоких широт Южного полушария, относящиеся к «сумчатым» морским ежам с развитой заботой о потомстве (см. ниже), хотя в этом семействе преобладают виды, не заботящиеся о потомстве.
Авторы сопоставили полученные данные с биологическими и биогеографическими характеристиками изученных видов. Биологических характеристик было шесть: размер взрослой особи, масса тела, максимальная продолжительность жизни, подвижность (расселительная способность) взрослых особей, плодовитость и размер «пропагулы» (то есть той стадии жизненного цикла, на которой животное покидает родителей и переходит к самостоятельной жизни: у кого-то это маленькая икринка, у кого-то — почти взрослая, тщательно выкормленная и выпестованная родителями молодая особь).
Никакой корреляции генетического полиморфизма с биогеографическими и экологическими показателями (площадь ареала, приуроченность к широтным зонам, водный или наземный образ жизни и т. п.) обнаружить не удалось (хотя биогеографические характеристики, надо признать, оценивались весьма грубо). Напротив, все шесть биологических характеристик достоверно коррелируют с полиморфизмом, в совокупности объясняя 73% вариабельности видов по показателю πs. Наилучшим предиктором полиморфизма, намного превосходящим в этом отношении остальные пять переменных, оказался размер пропагулы (рис. 2).
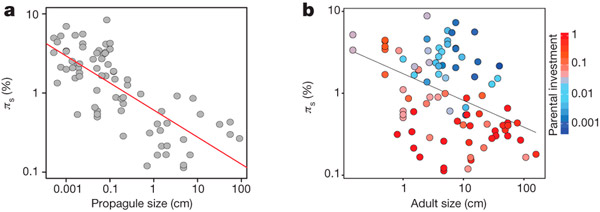
Рис. 2. Размер пропагулы (a, Propagule size) коррелирует с уровнем генетического полиморфизма (πs) гораздо сильнее, чем размер взрослого животного (b, Adult size). При этом даже та слабая корреляция, которая все же наблюдается во втором случае, в основном сводится к тому, что для крупных животных в среднем характерен больший родительский вклад в потомство, чем для мелких. Родительский вклад (Parental investment), определяемый как отношение размера пропагулы к размеру взрослой особи, обозначен на рисунке b оттенками красного и синего (см. шкалу справа). Рисунок из обсуждаемой статьи в Nature
В этом и состоит главная закономерность, обнаруженная авторами: чем более крупных потомков выпускают родители в мир, тем ниже (в среднем) генетический полиморфизм вида. Размер пропагулы, в свою очередь, отрицательно коррелирует с плодовитостью, причем эта корреляция весьма сильна. Таким образом, низкий полиморфизм характерен для животных, производящих небольшое количество тщательно выпестованных, хорошо подготовленных к самостоятельной жизни потомков (K-стратегия; см. Теория r-K отбора), а высокий — для тех, кто производит множество мелких и слабых потомков, бросая их на произвол судьбы (r-стратегия).
Размер взрослого животного коррелирует с полиморфизмом намного слабее (рис. 2, b). Этот результат является неожиданным, потому что размер, как правило, отрицательно коррелирует с численностью (популяции крупных животных в среднем имеют меньшую численность). Логично было бы предположить, что именно размер взрослого животного будет наилучшим предиктором полиморфизма, но это не подтвердилось. Среди видов с низким полиморфизмом есть как мелкие животные (менее 1 см), так и очень крупные. Виды одинакового размера могут иметь контрастно различающиеся уровни полиморфизма, если одни из этих видов являются K-стратегами, а другие — r-стратегами. Например, из рассмотренных авторами пяти видов морских ежей (Echinocardium cordatum, Echinocardium mediterraneum, Abatus cordatus, Abatus agassizi, Tripylus abatoides) первые два не заботятся о потомстве, производят много мелких яйцеклеток с небольшим количеством желтка, и поэтому их потомкам приходится начинать самостоятельную жизнь в виде крохотных, питающихся планктоном личинок — эхиноплутеусов. Три последних вида относятся к марсупиальным (сумчатым) морским ежам, чьи самки производят крупные, богатые желтком яйца и вынашивают молодь в специальных выводковых камерах, представляющих собой видоизмененные органы дыхания (петалоиды). У этих видов «пропагула», переходящая к самостоятельной жизни, представляет собой уже вполне сформированного морского ежика диаметром в несколько миллиметров. Соответственно, у первых двух видов полиморфизм высокий (πs = 0,0524 и 0,0210), у трех последних — низкий (0,0028, 0,0073, 0,0087). При этом по размеру взрослых особей все пять видов мало отличаются друг от друга. Аналогичная картина характерна для K- и r-стратегов среди офиур, немертин, насекомых и др.
Что касается насекомых, то в категорию K-стратегов здесь попали эусоциальные виды: термиты, пчелы, муравьи. В данном случае очевидно, что по размеру взрослой особи нельзя судить об Ne: численность (N) муравьев может быть очень высокой, что соответствует их небольшим размерам, но размножаться из них могут лишь немногие — «цари» и «царицы» (Ne << N). Понятно, что K-стратегия, обусловленная эусоциальностью, ведет к резкому снижению Ne. В других случаях этот эффект не столь очевиден.
Тем не менее авторы полагают, что обнаруженная связь между K-стратегией и низким полиморфизмом обусловлена именно отрицательным влиянием K-стратегии на эффективную численность, даже если природа этого влияния пока непонятна. Альтернативное объяснение могло бы состоять в том, что для K-стратегов характерен пониженный темп мутагенеза. Однако факты говорят скорее об обратном: темп мутагенеза (среднее число мутаций на геном за поколение), судя по всему, у K-стратегов выше, чем у r-стратегов. Одна из причин в том, что K-стратеги живут в среднем дольше, а поколения у них разделены большим числом клеточных делений (см.: У шимпанзе, как и у людей, число мутаций у потомства зависит от возраста отца, «Элементы», 18.06.2014). Так что различия в темпах мутагенеза скорее должны ослаблять, чем усиливать найденную отрицательную корреляцию между вкладом в потомство и полиморфизмом.

Рис. 3. Попарные корреляции между исследованными показателями: piS — синонимичный полиморфизм, piN — несинонимичный полиморфизм, piNpiS — отношение второго к первому, PropaguleSize — размер пропагулы, Longevity — продолжительность жизни, Fecundity — плодовитость, Size — размер, BodyMass — масса тела, Speed — скорость (подвижность). Цвета отражают величину коэффициента корреляции Пирсона (см. шкалу справа). Рисунок из обсуждаемой статьи в Nature
Уровень несинонимичного (значимого) нуклеотидного полиморфизма (πn) у исследованных видов тоже лучше всего коррелирует с размером пропагулы, хотя эта корреляция слабее, чем для синонимичного полиморфизма (рис. 3). Отношение πn/πs широко варьирует у разных видов и сильнее всего коррелирует с продолжительностью жизни: у долгоживущих организмов доля несинонимичных полиморфизмов повышена. Этот результат легко объясним: у долгоживущих видов, при прочих равных условиях, эффективная численность популяции должна быть ниже, а дрейф сильнее. Следовательно, слабовредные значимые мутации у долгожителей отбраковываются менее эффективно.
Таким образом, исследование показало, что уровень генетического полиморфизма можно довольно точно предсказать, зная определенные аспекты биологии рассматриваемого вида, такие как величина родительского вклада в потомство, приверженность K- или r-стратегии и продолжительность жизни. Случайные колебания численности популяции, полагавшиеся до сих пор чуть ли не главным фактором, влияющим на уровень полиморфизма, по-видимому, играют менее важную роль в глобальном масштабе. Хотя, конечно, нельзя отрицать их определяющее значение во многих частных ситуациях (как у тех же гепардов).
Авторы полагают, что K-стратегия в долгосрочной перспективе должна коррелировать с более низкими значениями Ne, а r-стратегия — с более высокими. Возможно, дело в том, что K-стратеги, благодаря эффективной заботе о потомстве, в целом более толерантны к низкой численности популяции: они могут долго существовать при низкой численности, не вымирая. Они могут, подобно гепардам, успешно восстановиться даже после экстремальных снижений численности, когда от всего вида остается несколько десятков особей. Напротив, r-стратеги сильнее зависят от изменений среды, которые приводят к резким колебаниям численности; их стратегия более «рискованная», поэтому в долгосрочной перспективе будут сохраняться только те виды, чья численность лишь очень редко или никогда не снижается до экстремально низких значений. Косвенно это рассуждение подтверждается палеонтологическими данными: во время массовых вымираний K-стратеги, по-видимому, имеют больше шансов выжить, чем r-стратеги, особенно крупноразмерные. Например, во время великого вымирания на рубеже мела и палеогена (66 млн лет назад) вымерли динозавры, у которых были большие проблемы с заботой о потомстве, — но выжили птицы и млекопитающие (ярко выраженные K-стратеги); вымерли аммониты (r-стратеги), но выжили наутилоидеи с крупными «пропагулами».
Исследование также заставляет задуматься об общих тенденциях эволюции K- и r-стратегов. На первый взгляд кажется, что у первых эволюционные перспективы должны быть в целом гораздо хуже, чем у вторых. У K-стратегов ниже смертность, особенно на ранних стадиях развития, что ограничивает возможности для естественного отбора. Как мы теперь знаем, у них ниже также и генетический полиморфизм, который считается важнейшим показателем «генетического благополучия» вида, приспособляемости и эволюционной пластичности. У них, вероятно, в среднем ниже и эффективная численность популяции. Это способствует ослаблению отбора и усилению дрейфа, что, в свою очередь, должно замедлять отбраковку слабовредных мутаций и фиксацию слабополезных. Стало быть, у K-стратегов должен быть большой мутационный груз (см. Genetic load). Если посмотреть на ситуацию под таким углом, то становится вообще непонятно, почему K-стратеги до сих пор не вытеснены повсеместно r-стратегами. В действительности, судя по палеонтологическим данным, тенденция скорее обратная, особенно у таких ключевых представителей наземной биоты, как сосудистые растения и четвероногие (наземные позвоночные). В этих группах в течение фанерозоя прослеживается явный сдвиг в сторону K-стратегии: беззащитные «пропагулы» в виде мельчайших спор и икринок уступают место увесистым семенам и подрощенным, хорошо упитанным детенышам.
По-видимому, K-стратегия каким-то образом компенсирует все вышеперечисленные недостатки. Одним из факторов может быть отмеченная выше толерантность к низкой численности, обусловленная меньшей зависимостью смертности от непредсказуемых колебаний среды: сравните положение беззащитных лягушачьих икринок в пруду и птичьих яиц в теплом гнездышке с заботливой наседкой. Кроме того, хотя у K-стратегов уровень смертности (элиминации) ниже, эта смертность наверняка более избирательна и неслучайна, чем у r-стратегов. Гибель мелких «пропагул» часто происходит случайно и вообще не зависит от качества генов. Возможно, отбор у K-стратегов даже при низком уровне смертности достаточно эффективен за счет более избирательной (зависящей от качества генов) элиминации. Наконец, можно допустить, что забота о потомстве делает многие потенциально вредные мутации (которые снизили бы шансы на выживание молодняка, брошенного на произвол судьбы) де-факто нейтральными. В этом случае часть значимого (несинонимичного) полиморфизма у K-стратегов в действительности может оказаться не «мутационным грузом» (слабовредными мутациями, не отбракованными своевременно из-за сильного дрейфа и слабого отбора), а нейтральным полиморфизмом, повышающим эволюционную пластичность.
Источник: J. Romiguier, P. Gayral, M. Ballenghien, A. Bernard, V. Cahais, A. Chenuil, Y. Chiari, R. Dernat, L. Duret, N. Faivre, E. Loire, J. M. Lourenco, B. Nabholz, C. Roux, G. Tsagkogeorga, A. A.-T. Weber, L. A. Weinert, K. Belkhir, N. Bierne, S. Glémin & N. Galtier. Comparative population genomics in animals uncovers the determinants of genetic diversity // Nature. 2014. V. 515. P. 261–263.
-
Известно, что К-стратеги хорошо переносят внезапные изменения среды и всяческие бутылочные горлышки в своей численности. Но вот интересно, что случается с K-стратегами, когда они наоборот испытывают нехарактерный всплеск численности на несколько порядков? Которую не так-то просто обуздать, и которая очень дорого даётся планете в силу огромных ресурсов, требующихся для поддержания такой численности, в силу как раз их К-стратегии: мало того, что особей много, так они еще и крупные, и живучие, и выхаживают своих детенышей до 21 года, и живут до 80-ти лет! И из-за этого они заселяют всю планету, и начинают влиять на всю экосистему вплоть до создания условий очередного вымирания планетарного масштаба... Есть ли что природе ответить на такой вызов?
-
Если верна теория РНК-мира, то в свое время на планете было огромное разнообразие "РНК", репликаторов. И разной хиральности. А в итоге остался весьма узкий набор, который обеспечили все разнообразие организмов , бывшее и существующее. В конце концов, если останется ограниченный набор видов, с человеком в основе, который эффективно сможет обеспечивать воспроизводство этих видов, то что с того? Не испытываю никакого ужаса по поводу того, что мяса и прочие вещи будут выращивать в специальных чанах, а не путем содержания коров.
-
1) Обуздать всплеск роста численности К-стратегов не сложно и даже непросто, а очень просто: достаточно ввести плотностнозависимую регуляцию численности. Тигр, не обзаведшийся своей территорией, избегает размножения, хотя кол-во жертв позволяет размножаться каждый год.
2)Человек, которого Вы привели в качестве примера, также подвержен плотностнозависимой регуляции - см. книгу С.П.Капицы "Парадоксы роста". В настоящий момент Европа прошла точку перегиба кривых роста, а Африка и Азия к ней приближаются. Ближайшие 200 лет все решат - наша собственная природа ответит на наш вызов.
3)Создание человеческой цивилизацией условий для очередного вымирания планетарного масштаба связано не с ее высокой численностью, а с высоким уровнем роскоши и алчности. Один легковой автомобиль на 6 пассажиров, в котором ездит максимум два человека, гадит природе больше, чем две сотни африканцев, занимающихся традиционным сельским хозяйством. Одноразовые пакеты, выбрасываемые странами большой восьмерки создают для природы больше проблем, чем все страны Африки с их высоким приростом населения. А вот алчность, в отличие от численности ничем не регулируема. Тут на вызов отвечать нечем.
4) А.Маркову следует следить за речью - нейтральные гены на то и нейтральны, что не могут входить ни в генетический груз, ни в мобилизационный резерв Шмальгаузена, определяющий пластичность организма. Фигура речи понятна. но некорректна.-
>Один легковой автомобиль на 6 пассажиров, в котором ездит максимум два человека
Помогает предотвратить очередной ледниковый период. :~]-
А негритянские коровы не помогают? Вместо выбросов CO2, которые неизвестно на что влияют (и влияют ли вообще - еще один сферический конь в вакууме) лучше подсчитайте конкретную - в квадратных километрах - площадь территории, которую отчуждает у биосферы один автомобиль за время своего существования и еще некоторое время после него. Причем считайте с обслуживающей автомобиль инфраструктурой. Зная Ваше пристрастие к сферическим коням в вакууме, напоминаю, что автомобиль обслуживает ВСЯ дорога на всем ее протяжении безотносительно того, сколько автомобилей по ней ездят и какое расстояние проезжают.
-
>напоминаю, что автомобиль обслуживает ВСЯ дорога на всем ее протяжении безотносительно того, сколько автомобилей по ней ездят и какое расстояние проезжают.
Т.е. чем их больше, тем меньше эта самая удельная площадь отчуждаемой территории на каждый автомобиль. Поэтому нужно стремиться к максимальной загруженности уже построенных автомагистралей! ;~] -
-
-
-
-
-
>Которую не так-то просто обуздать
Дык уже - демографический переход даже в китаях и индиях подходит к концу (как тут уже выше заметили).
А по оценкам, что нам встречались - так до предела ещё в 1,5 раза можно расти.
>и которая очень дорого даётся планете в силу огромных ресурсов, требующихся для поддержания такой численности
Насколько дорого в граммах? Плейстоценовой мегафауной? Ну и черт бы с ней. И вообще тогда растущие человеки ничем больше питаться кагбе особо и не умели. А в будущем может даже и получится частично исправить "ошибки молодости" - вон плейстоценовый парк уже сейчас пытаются делать.
А за удвоение выбросов углекислого газа и азотные удобрения биосфера вообще должна спасибо сказать за предотвращение очередного ледникового периода и за повышение продуктивности путем многократного увеличения азотфиксации.
Такие дела.


Появление "смысла смерти" - это увеличение сложности организации вида. Чем больше выживаемость индивида зависит от случайных факторов среды - тем ниже организация вида, тем более он зависим от "воли богов", тем более он является продуктом среды. Чем больше выживаемость индивида зависит от кода в его ДНК, тем более высокоорганизован вид, тем более его генофонд "чувствует" механику среды. И тем важнее индивиду "правильно умереть" (тем конкретнее в его ДНК "записана" его "судьба").
(Эдакая кибернетическая формализация биологической эволюции: превращение внешних сигналов во внутренние обратные связи, в "предсказания" "генетического оракула".)
-
1) А причем здесь смерть, если "cмертный бой не ради славы, ради ЖИЗНИ на Земле"? Вставьте в Ваш пост "жизнь" вместо "cмерть" и его смысл не изменится.
2)"Чем больше выживаемость индивида зависит от кода в его ДНК" - тем более он привязан к окружению. Вы рисуете мещанина-приспособленца, которому где хорошо, там и Родина. Мне больше нравится Нитцше - сверчеловек сам определяет когда ему умереть, а когда жить. При этом сверхчеловек обязан делать добро, но не из сострадания, а под угрозой утраты статуса сверхчеловека.-
1) Согласен, что жизнь и смерть в моей формулировке практически одно и то же. Однако заменить смерть на жизнь можно только после смерти, когда "все показания" с жизни индивида уже сняты "генетическим оракулом" (гарантируется, что он больше не участвует в динамике генофонда, в популяции с культурной эволюцией это уже не так - там помимо генов начинают жить мемы, и какой-нибудь аристотель может влиять на умы людей даже после своей смерти).
2) Я ни в коем случае не определяю смысл жизни человека, разумного существа. Я говорю лишь об биологической эволюции. А насколько ею детерминирована судьба человека - вопрос открытый (и я тоже склонен выбирать вариант свободного духа, порождённого, но не связанного природой). Так уж сложилось, что даже самые великие умы состоят из мещан-приспособленцев - клеток.-
Вы увлекаетесь ролью разума ибо
1) "Показания" с жизни индивида cнимаются и после его смерти. Дрозофила дрозофилу не помнит, но собака собаку помнит очень долго. У слонов в неволе память - более 20 лет. Мертвое дерево, стоящее на корню, продолжает влиять на окружающую экосистему пока стоит (а упав, продолжает влиять, пока не сгниет). А влияние организмов-рифообразователей вообще растягиваются на миллионы лет после их смерти. Как видите, не нужно большого ума, чтобы оставить по себе память. Вместо большого ума достаточно быть просто большим.
2)"Я ни в коем случае не определяю смысл жизни человека, разумного существа. Я говорю лишь об биологической эволюции." - Хотите cказать, что животные не понимают, что рискуют? Что верблюд, уходя от оазиса не отдает себе отчета или белый медведь, решивший доплыть до берега 30 км. не понимает, чем это ему грозит? Пример холодных зимовок уток в истоке Ангары наглядно демонстрируют насколько адекватно птица воспринимает риск перелета через холодный Байкал. Хотя противоположный берег виден невооруженным глазом, а утка - птица водоплавающая.
3)А Нитцше тоже не определяет жизнь разумного человека. Сверхчеловек выше разума.
4) "Так уж сложилось, что даже самые великие умы состоят из мещан-приспособленцев - клеток" - не примите, пожалуйста за грубость или сарказм, но так уж сложилось, что чем проще предрассудок, тем он устойчивее. 30 лет назад Черданцев показал, что клетки зародыша нельзя рассматривать как мещан-приспособленцев. К популяции зародышевых клеток приложима теорема Фишера, но не дилемма Холдейна, а значит применять понятия генетического груза и генетического разнообразия (равно как и нейтрального признака) здесь некорректно.Если уж кто приспособленец, то это многоклеточная морфологическая структура, формирующаяся последней из зародышевой закладки. Или раковая клетка. Но из нее не просто великий, из нее вообще никакого ума не построишь.Иначе раковые клетки формировали бы свои собственные виды. Но такой случаев зафиксировано очень мало: трансмиссивная венерическая саркома собак и лицевая саркома сумчатого дьявола.-
Думаю, вы правильно увидели слабые стороны моей "недомодели", но вообще акцент был на другое. "Генетический оракул" (по аналогии со "случайным оракулом") - вот концепция, позволяющая смоделировать влияние внешних и внутренних сигналов на генофонд (в том числе "постсмертных" - набор типов сигналов можно подбирать экспериментально). Получится более обобщённый генетический алгоритм, в котором выразимы не только свойства (гены) искомых решений, но и процессы их взаимодействия (т.е., возможен результат, не являющийся следствием полового процесса, это может быть любой абстрактный процесс, последовательность сигналов с определённым распределением вероятностей). При этом "смерть" в этой модели - важный тип "сигнала", сильно изменяющий вероятность последующих сигналов (даже если таковые и вводятся в модель).
В общем, я согласен, что слишком сумбурно озвучил какую-то мутную концепцию. Это просто ассоциации фактов из прочитанной статьи с моими наработками в теме эвристических алгоритмов. Возможно, эти аналогии слишком ограничены или даже некорректны.
А с четвёртым пунктом не соглашусь - ваши интерпретации мне понятны, но более удобной кажется докинзова концепция эгоистичного гена. Смерть индивида в ней тоже можно считать "мещанством" (как и смерть клетки, и даже вида, но в разных степенях), направленным на сохранение (увеличение вероятности появления в будущем) генов этого индивида, т.к. генофонду в этом году "выгодно" такое же количество смертей, как и в прошлом, отклонения в любую сторону для него "болезненны" (это очень грубо, на самом деле возможны циклические и ациклические колебания "оптимума" или монотонные тренды и, конечно, смерть без потомства чаще всего менее "желательна" с точки зрения генофонда, но далеко не всегда). Каждый из генов сам по себе не конкретный объект, а некое облако вероятностей возникновения в реальности в сочетании с другими генами, которые рекурсивно так же определяются такими же вероятностными облаками (потому модель позволяет и на уровне клеток, и на уровне видов воспроизводить динамику - для неё это просто разный набор вероятностей). В общем-то, "генетический оракул" (название придумалось в предыдущих комментариях) - это и есть попытка математического описания динамики таких вот объектов.
Сигналы-аналоги "ухаживания родителей" (например, передача "энергии" родителя своему ребёнку, даже с существенными потерями) в моей модели тоже приводят к снижению "видового полиморфизма", но увеличивают сложность "поведения" отдельного "индивида" (тенденция "энергии" "протаптывать" себе более-менее статичные "русла протекания" по последовательностям типов событий, даже если для этого необходимо усложнение этой последовательности).
А отдельные гены, набравшие очень большие вероятности сочетаний с ограниченным набором генов в ходе моделирования претендуют, наверное, на роль вирусных частиц (хотя сам механизм отрыва от генофонда и горизонтальный перенос генов пока не моделируется - нужно вводить новые типы сигналов). Или должны стать видообразующим фактором (возникновение репродуктивной изоляции тоже пока не моделируется).-
Если не соглашаетесь с пунктом 4, то как в Вашей модели (равно как и в докинзовой) гены вообще общаются со средой? Нормальный организм общается благодаря набору рецепторов, причем он на то и организм, чтобы отличать хорошее от плохого. Ни один ген на это не способен (прошу прощения, но я имею ввиду реальные гены, а не генноинженерные конструкции). Ergo любой ген будет эволюционировать тогда и только тогда, когда это разрешит организм. У обезьяны появился ген, увеличивающий размер мозга. Думаете, такая обезьяна получит преимущество (поклонники эгоистичного гена так и думают)? Как бы не так - маленькая тиреойдная железа не сможет обслужить развитие большого мозга и вместо великого гения родится несчастный кретин (но зато с великолепной генетикой!). Разрешение бывает и чисто волевым. Допустим, у верблюда появился ген, благоприятствующий тебеневке (выкапыванию корма из под снега). Но ни один верблюд не выкапывает корма из под снега и вообще избегает заснеженных мест, даже если с ними встречается (например, в Южной Сибири и Казахстане). Верблюду глубокий снег элементарно неприятен, а уж для того, чтобы копаться в нем необходимо волевое усилие. Не будет усилия (а верблюды - редкие упрямцы, говорю как бывший работник зоопарка), не будет и эволюции даже если нужные гены есть. Так что, для эволюции нужен сверхчеловек Нитцше (или хотя бы сверхверблюд:-))/ Эгоистичному гену не менее эгоистичный организм не даст эволюционировать.
-
-
-
-
Если так - у бесполых(у которых геном потомка тождественен геному предка безотносительно уровня полиморфизма) описанной зависимости быть не должно.
-
Четыре делится на два, потому что это чётное число, или четыре - чётное число, потому что делится на два?
-
Математика - не наука, там как условятся - так и есть,
а в биологии таки либо K-стратегия снижает полиморфизм, либо высокий полиморфизм способствует отказу от К-стратегии, либо оба фактора зависят от одного и того-же третьего.-
Математика - наука, и поесстественней традиционно признаваемых естественными. Ваша ди(три-)хотомия зависит от аксиом, совокупность коих названа "биологией". В этой системе и "К-стратегия" и "полиморфизм" являются абстракциями, выведенными из ограниченных данных. Притом "вывод" означает произвольное применение правил логики, бритвы Оккама и прочих надавливаний коленкой. В таких условиях абсолютизация абстракций вредна.
-
1) Не надо путать. Математика - не естественная, а точная наука. Точные науки - математика, физика, частично химия - имеют дело с самоидентичными объектами. Число 3 всегда число 3, получили ли его вычитанием, делением или тройным интегрированием, вчера или 200 лет назад. Атом хлора (прошу не путать с изотопом) всегда атом хлора, неважно каким способом и когда он был получен. Они идентичны себе и, соответственно, всем тройкам и атомам хлора, какие есть во всех существующих Вселенных, равно как и в тех, которые существовали и будут существовать. А вот любое живое существо несамоидентично даже в нашей Вселенной: на сытой акуле можно верхом прокатиться, спящую (точнее - в оцепенении) за хвост подергать, только что родившуюся - в аквариуме держать. А ведь это одна и та же акула, причем даже не вид, а особь. И я не завидую тому математику, который встретит в океане эту особь как голодную, взрослую и бодрствующую. В ее челюстях он может и поймет разницу между точными и естественными науками, между самоидентичными и несамоидентичными объектами. Но уже никому об этом не расскажет.
2) "Притом "вывод" означает произвольное применение правил логики, бритвы Оккама и прочих надавливаний коленкой". - это в точных науках. В естественных все вышеперечисленное ограничивается классификацией. Причем, ввиду несамоидентичности, постороение классификации всегда априорно (точнее - интуитивно-априорно, человек ведь тоже живое существо и поэтому может обращаться к собственноым ощущениям; антропоморфизм - первая естественнонаучная классификация), с последующим улучшением по принципу вероятностного прогнозирования. Точные науки, в отличие от естественных, никогда не строят априорные классификации. Впрочем, чаще они прекрасно обходятся вообще без классификаций. Арифметику создали без теории чисел, ньютонову механику - без теории пространства. Создать биологию без априорно-интуитивной классификации (хотя бы на уровне живое-неживое, причем, заметьте - это самый высокий из всех возможных в биологии уровней) невозможно. Точные науки двигаются от простого к сложному, а естественные - от сложного к простому.-
Традиционно говоря, это неестественная наука. Физика, химия, биология - естественные. Есть, как все знают, еще противоестественные. "Самоидентичность" - это абстракция. Акулой называют конкретную особь в конкретном состоянии, имея в виду ее абстракный обобщенный образ. Это типичный акт абстрагирования, необходимый для применения логики. Ваша априорная классификация - из той же оперы, и без бритвы Оккама, отсекающей ненужные параметры вроде сытости акулы, сколько-нибудь удобную классификацию построить нельзя. Про арифметику и механику и ненужность для них априорных классификаций - это абсурд! (Очень смешно). Арифметика и есть теория чисел, одна из простейших, а ньютонова механика и есть теория пространства, одна из не самых сложных. Все без исключения науки, если это науки, начинаются с априорной классификации, которая является составной частью соответствующей аксиоматической системы. В арифметике числа, действия с ними и обобщение понятия на результат операций - по большей части априорная классификация, позволяющая ввести удобную аксиоматику.
-
1) "Самоидентичность" - это абстракция. А атом хлора тоже абстракция?
2) "Акулой называют конкретную особь в конкретном состоянии, имея в виду ее абстракный обобщенный образ" - ерунду говорите. Сразу видно, с бинарным ключом не работали, ни одного растения по числу тычинок и форме пестика не определяли. Не говоря уже об акуле. Ламну от кархарины отличают не по "ее абстракному обобщенному образу", а по конкретным зубам (и часто кроме зубов ничего нет).
3) "и без бритвы Оккама, отсекающей ненужные параметры вроде сытости акулы, сколько-нибудь удобную классификацию построить нельзя" - Снова говорите ерунду. Бинарный ключ Ламарка, с которым каждого биолога знакомят на первом курсе университета (как правило, стыдливо замалчивая кто именно эту великую штуку придумал), построен именно без бритвы Оккама. Но самый выдающийся пример - это таблица Менделеева. Ключ Ламарка - это все-таки способ упорядочивания наших представлений о мире, а таблица Менделеева - это сам мир. Французские химики, самые позитивистки подкованные в то время, его за пренебрежение "типичным актом абстрагирования" очень ругали, считая атомный вес именно ненужным абстрактным понятием. Говорили Вашими словами - это абсурд! Действительно, в то время атомный вес считали, усредняя данные по всем месторождениям минералов, подозреваемых на данный элемент (кстати, именно поэтому все учебники врут,если пишут, что до Менделеева атомный вес такого-то и такого-то элемента был определен неправильно. Еще как правильно! Именно так и нужно определять с точки зрения поклонников "типичного акта абстрагирования"). Поклонники бритвы Оккама и типичного акта абстрагирования предрекали, что вся таблица Менделеева полетит к чертям после открытия новых месторождений. Менделеев тоже до конца не изжил в себе преклонение перед бритвой Оккама, поэтому так и не опубликовал самое большое предвидение своей таблицы - группу одноатомных тяжелых элементов с низкой реакционной способностью. Теперь их называют инертными газами. И не понял открытия Аррениуса, считая ионы лишними сущностями. Так и говорил - недоатомов не бывает, Аррениус - лжеученый.
4) "В арифметике числа, действия с ними и обобщение понятия на результат операций - по большей части априорная классификация", - откройте тайну, а что тогда находится "по меньшей части"?
5) "а ньютонова механика и есть теория пространства, одна из не самых сложных" - Вы путайте Ваше представление о механике Ньютона и представление самого Ньютона. Ошибка типичная. Надо полагать, с теорией парадигм Куна Вы знакомы очень поверхностно. Почитайте.
6) "Все без исключения науки, если это науки, начинаются с априорной классификации, которая является составной частью соответствующей аксиоматической системы". - поздравляю с "открытием" в науковедении. Еще раз советую почитать книгу Куна. Тогда узнаете, что все науки начинаются со списка разрешенных задач и образцов их решения. К классификации это не имеет никакого отношения.-
Ваша позиция в этой теме мне более не интересна. Не то, чтобы она была неверной, просто это художественная литература. Отвечу только про "меньшую часть" - это изобретение новых сущностей из соображений красоты.
-
-
-
-
-
-
-
>Докажите, что организм отличает родственника от неродственника, чтобы оказать родственнику помощь.
По моему, это как раз те, кто считает, что К-стратегия приводит к уменьшению полиморфизма должны что-то подобное доказать (и кстати, характерная для К-стратегов более тщательная оценка партнёра для спаривания, наверно, действительно может приводить к определённому снижению полиморфизма...)
Мне же кажется, что высокий уровень полиморфизма должен снижать эффективность всех видов родственного отбора, неэффективность отбора приводит к вырождению признаков, отвечающих за реализацию К-стратегии(хотя признаки отнюдь не становятся неважными, нарушается именно работа отбора), в результате чего вид либо становится эффективным r-стратегом, либо просто неэффективным К-стратегом - что приводит к снижению численности и уменьшению полиморфизма.-
1) С чего Вы взяли, что именно К-стратеги более тщательно оценивают партнёра для спаривания? Загляните сюда, чтобы убедиться в обратном http://lenta.ru/news/2014/11/18/sealspenguins/. Роль полового отбора сильно преувеличена сторонниками концепции эгоистичного гена.
2) "Мне же кажется, что высокий уровень полиморфизма должен снижать эффективность всех видов родственного отбора" - согласен с Вами с поправкой - любого отбора, если рост разнообразия не подкреплен ростом численности. Дилемма Холдейна.
-
Последние новости










Рис. 1. Уровень генетического разнообразия у 76 видов животных. Слева — эволюционное дерево, отражающее родственные связи изученных видов. Эти виды относятся к 31 семейству (для каждого семейства приведено схематичное изображение одного из представителей) и представляют 8 типов животных (названия типов приведены справа). Левая разноцветная колонка отражает уровень нейтрального генетического полиморфизма для каждого вида (см. шкалу вверху); πs — процент синонимичных сайтов, различающихся у двух случайно выбранных гомологичных последовательностей (мера нейтрального полиморфизма). Средняя разноцветная колонка отражает размер «пропагулы», то есть организма на той стадии развития, когда он покидает родителей и переходит к самостоятельной жизни (мера родительского вклада). Правая колонка характеризует географическое распространение вида. Рисунок из обсуждаемой статьи в Nature