Изменение окраски помогает цветам разделиться на два вида даже без смены насекомого-опылителя

Два вида флоксов, Phlox drummondii и P. cuspidata, имеют одинаковые бледно-голубые цветки в непересекающихся (аллопатрических) частях своих ареалов, но в симпатрической части, где оба вида встречаются вместе, у первого из них преобладают ярко-красные цветки. Эксперименты, проведенные американскими биологами, показали, что измененная окраска является адаптацией для предотвращения межвидовых скрещиваний. Хотя оба вида опыляются одними и теми же видами бабочек, каждая отдельная бабочка, как правило, предпочитает либо яркие, либо бледные цветки. Поскольку межвидовые гибриды отличаются пониженной плодовитостью, такое поведение опылителей приводит к тому, что растения P. drummondi с ярко-красными цветами в симпатрической зоне размножаются эффективнее, чем представители того же вида с голубыми цветами. Исследование продемонстрировало в действии один из классических механизмов видообразования: отбор, обусловленный пониженной приспособленностью гибридов, приводит к выработке адаптаций, препятствующих гибридизации.
Согласно одной из классических эволюционных моделей, формирование репродуктивной изоляции между двумя зарождающимися видами может происходить в два этапа. Сначала по тем или иным причинам (например, из-за проживания на разных территориях) между двумя частями предкового вида накапливаются генетические различия. Некоторые из них непременно будут взаимно несовместимыми — просто потому, что отбор не «проверяет» изменения, возникающие в двух разобщенных популяциях, на совместимость друг с другом. Поэтому рано или поздно возникает частичная постзиготическая изоляция. Это значит, что гибридное потомство будет теперь иметь пониженную приспособленность по сравнению с «чистыми» представителями двух популяций. Об этом механизме видообразования, известном как «модель Добжанского-Мёллера», рассказано в заметке «Генетическая несовместимость нарастает по параболе» («Элементы», 26.09.2010).
Впоследствии представители двух разобщенных популяций, между которыми уже возникла частичная постзиготическая несовместимость, могут встретиться и получить возможность скрещиваться друг с другом. Например, такое может произойти при исчезновении физико-географических барьеров, ранее разделявших области их распространения. В этом случае должен вступить в действие так называемый механизм усиления (reinforcement). Суть его в том, что отбор, обусловленный пониженной приспособленностью гибридов, будет поддерживать любые мутации, способствующие презиготической изоляции, то есть снижению вероятности гибридизации.
В последние годы механизм усиления активно изучается, и на сегодняшний день уже понятно, что это явление широко распространено в природе. В большинстве случаев, однако, о нем приходится судить по косвенным данным (см. Новые виды образуются очень быстро, если меняется система распознавания половых партнеров, «Элементы», 30.04.2010; О том, как встреча после разлуки завершает видообразование, синопсис Елены Бадьевой по статье В. А. Лухтанова «Правило Добржанского и видообразование путем усиления презиготической репродуктивной изоляции в зоне вторичного контакта популяций», ЖОБ, том 71, 2010. № 5, Сентябрь–октябрь. Стр. 372–385).
Два вида флоксов, растущих в Северной Америке, — Phlox drummondii и P. cuspidata — судя по косвенным признакам, являют собой типичный пример механизма усиления в действии. В аллопатрических (непересекающихся) частях ареалов цветы у них одинакового бледно-голубого цвета. Такая окраска является исходной (анцестральной) для флоксов. Однако в симпатрической зоне — в восточном Техасе, где ареалы двух видов пересекаются, — среди представителей P. drummondii преобладают растения с ярко-красными цветами. Межвидовая гибридизация у этих видов «технически» вполне возможна, и она действительно происходит в природе. Гибриды, однако, отличаются пониженной приспособленностью: их плодовитость на 90% ниже, чем у «чистых» представителей обоих видов. Следовательно, флоксам крайне невыгодно скрещиваться с «чужаками». Поэтому логично предположить, что изменение окраски цветов у P. drummondii с бледно-голубой на ярко-красную — это адаптация для предотвращения межвидовой гибридизации. Остается лишь доказать, что это действительно так, и понять, каким образом окраска цветов препятствует межвидовым скрещиваниям.
Именно эти две задачи и удалось решить Робину Хопкинсу (Robin Hopkins) и Марку Раушеру (Mark D. Rausher) из Университета Дьюка (США).
Генетические основы различий по окраске цветов у P. drummondii известны. Различия определяются двумя генами, каждый из которых существует в виде двух вариантов (аллелей). Первый ген (F3’5’h) кодирует фермент (флавоноид-3’5’-гидроксилазу), участвующий в синтезе пигментов антоцианов. Исходный (анцестральный, предковый) аллель этого гена (H) обеспечивает синюю или голубую окраску цветов. Производный аллель (h) возник в результате мутации в регуляторной области гена, которая привела к снижению его экспрессии, что, в свою очередь, изменило оттенок лепестков с голубого или синего на розовый или красный. Аллель H является доминантным по отношению к h, то есть, растения с генотипами HH и Hh имеют синие или голубые цветы, растения с генотипом hh — красные или розовые. Этот ген можно назвать «геном оттенка».
Второй ген (R2R3-Myb) можно назвать геном интенсивности или яркости окраски. Он кодирует транскрипционный фактор, от которого зависит количество производимых пигментов и, соответственно, интенсивность окраски цветов. Исходный аллель i обеспечивает бледную окраску (голубую или розовую), производный аллель I придает цветам ярко-синий или ярко-красный цвет. Аллель I доминантен по отношению к i.
В аллопатрической части ареала P. drummondii все растения имеют генотип iiHH и, соответственно, голубые цветы. В симпатрической части подавляющее большинство растений этого вида имеют генотип IIhh и ярко-красные цветы. «Рекомбинантные» растения iihh с розовыми цветами и I-H- с ярко-синими встречаются только на границе симпатрической и аллопатрической частей ареала.
Чтобы проверить гипотезу о том, что ярко-красная окраска цветов в симпатрической области является адаптацией для предотвращения межвидовой гибридизации, прежде всего нужно было убедиться, что эта окраска не дает флоксам каких-то иных преимуществ. Ведь могло оказаться и так, что природные условия восточного Техаса почему-то благоприятствуют красным цветам у данного вида флоксов, а механизм усиления тут ни при чем.
Авторы вывели четыре линии флоксов P. drummondii, гомозиготных по генам окраски (IIHH — синие, IIhh — красные, iiHH — голубые, iihh — розовые) с рандомизированным «генетическим фоном». Семена всех четырех типов затем высаживались в равной пропорции на огражденных участках на территории биостанции Техасского университета (Stengl Biological Station). Биостанция расположена в зоне пересечения ареалов двух видов флоксов. Оба вида встречаются в диком состоянии на территории биостанции, но перелеты бабочек-опылителей с диких цветов на опытные участки были пренебрежимо редким событием по сравнению с перелетами с цветка на цветок в пределах участка.

Схема выведения четырех подопытных линий P. drummondii. Скрещивали дикие голубые цветы с дикими ярко-красными (Parent), потомков первого поколения (F1) размножали самоопылением, затем скрещивали растения второго поколения (F2) с одинаковой окраской, но из разных линий, чтобы получить четыре гомозиготных линии с рандомизированным генетическим фоном (Experimental seeds). Изображение из дополнительных материалов к обсуждаемой статье в Science
Оказалось, что при отсутствии риска межвидовой гибридизации ни выживаемость, ни плодовитость флоксов P. drummondii не зависят от окраски цветов. Все четыре генотипа продемонстрировали одинаковый уровень приспособленности. Это — весомый аргумент в пользу того, что ярко-красная окраска не является адаптацией к местным условиям среды.
Эксперименты по искусственному опылению показали, что никакой защиты от межвидовой гибридизации на уровне взаимодействия пыльцы и пестика у изучаемых флоксов нет: оплодотворение при внутри- и межвидовых скрещиваниях происходит одинаково успешно.
Теперь нужно было проверить, помогает ли ярко-красная окраска избежать межвидовой гибридизации. Для этого на каждом участке сажали в случайном порядке 30 саженцев одной из четырех гомозиготных линий P. drummondii, 105 контрольных саженцев голубых P. drummondii из аллопатрической части ареала и 115 флоксов другого вида — P. cuspidata (у которых цветы всегда голубые). Когда растения вырастали и давали семена, авторы брали по 100–150 случайно выбранных семян от опытных и контрольных растений P. drummondii и при помощи генетического анализа устанавливали их «отцовство», подсчитывая число гибридных семян в каждой выборке.
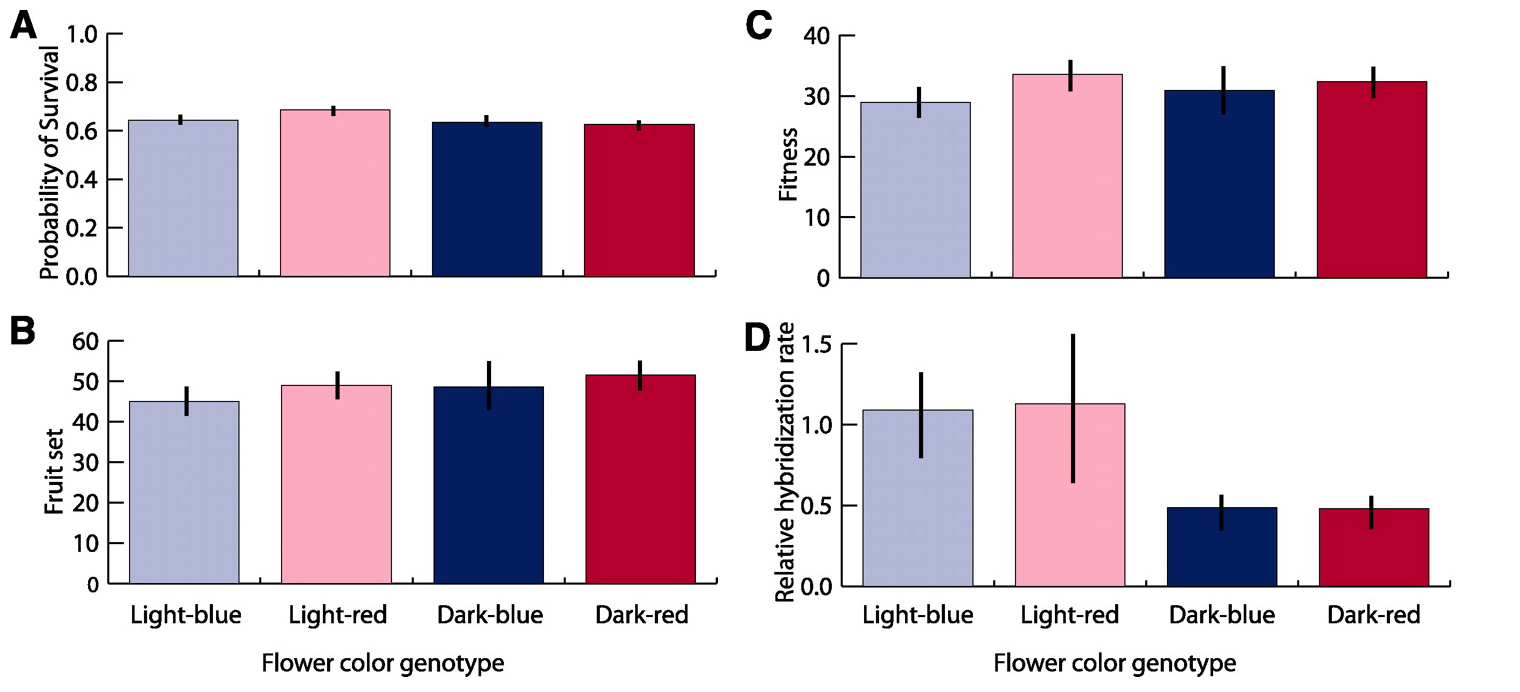
Жизненные характеристики флоксов P. drummondii с разной окраской цветов. A — выживаемость, B — плодовитость (количество плодов), C — «общая приспособленность» (произведение выживаемости на плодовитость), D — относительная частота межвидовой гибридизации при совместном произрастании с флоксами другого вида (P. cuspidata), по сравнению с контрольными растениями с голубыми цветами из аллопатрической части ареала P. drummondii. Изображение из обсуждаемой статьи в Science
У контрольных голубых растений доля гибридных семян оказалась весьма высокой: от 28 до 44% на разных участках. Столь же высокий уровень межвидовой гибридизации был обнаружен у гомозиготных голубых и розовых P. drummondii. Однако у ярко-красных и ярко-синих флоксов доля гибридных семян оказалась вдвое ниже. Таким образом, яркость окраски (но не ее оттенок) эффективно защищает флоксы от межвидовых скрещиваний. Поскольку гибридные растения имеют резко пониженную плодовитость, из этого следует, что там, где флоксы двух видов растут рядом друг с другом, действует сильный отбор, благоприятствующий аллелю яркой окраски I и обраковывающий растения с аллелем бледной окраски i.
Зная частоту гибридизации для разных генотипов и плодовитость гибридов и «чистых» представителей P. drummondii, можно рассчитать силу отбора, характеризуемую коэффициентом отбора s (см. Естественный отбор: количественная характеристика). В данном случае «яркий» аллель I поддерживается чрезвычайно сильным отбором (s=0,32), что должно вести к полному вытеснению «бледного» аллеля i в симпатрической зоне за считанные поколения.
Каким образом яркая окраска защищает от межвидовых скрещиваний? Ответить на этот вопрос помогли наблюдения за бабочками-опылителями. Главным опылителем обоих видов флоксов в симпатрической зоне является бабочка Battus philenor из семейства парусников (Papilionidae). Кроме того, в опылении участвуют различные виды бабочек-толстоголовок (семейство Hesperiidae). Как выяснилось, обе разновидности опылителей демонстрируют сходное поведение на участках, где разноцветные флоксы растут вперемешку. Бабочки не делают различий между голубыми и розовыми цветами. Например, насекомое, посетившее голубой цветок, в следующий раз с равной вероятностью сядет на голубой или на розовый. К какому виду относится растение с голубыми цветами (к P. drummondii или P. cuspidata), бабочкам тоже все равно. Этот факт хорошо согласуется с тем, что уровень межвидовой гибридизации одинаково высок у голубых и розовых P. drummondii.
Совсем другая картина наблюдалась на участках, где наряду с голубыми цветами росли ярко-красные или ярко-синие. Здесь бабочки проявляли значительное постоянство, совершая перелеты между цветами одинаковой яркости намного чаще, чем между яркими и бледными цветами. Так, бабочка, посетившая голубой цветок P. cuspidata, сядет после этого на голубой или розовый цветок P. drummondii с вдвое большей вероятностью, чем на ярко-синий или ярко-красный цветок. Этот факт полностью объясняет вдвое меньшую частоту межвидовой гибридизации у растений с яркими цветами. В обратную сторону закономерность тоже работает: если бабочка села на яркий цветок, после этого она, скорее всего, посетит другой яркий цветок, а не голубой или розовый. Следовательно, яркая окраска не только помогает избежать попадания пыльцы P. cuspidata на пестики P. drummondii, но и препятствует растрачиванию пыльцы второго вида на опыление «чужаков».
Полученные результаты показывают, что яркая окраска цветов в симпатрической зоне закрепилась у P. drummondii под действием механизма усиления как адаптация для предотвращения гибридизации. Эффективность этой адаптации обусловлена особенностями поведения бабочек-опылителей. Ранее уже было известно, что в пределах одного вида насекомых, питающихся нектаром, у разных особей могут быть разные предпочтения при выборе цветов. Эти различия могут быть отчасти врожденными (генетически обусловленными), отчасти — приобретенными, то есть связанными с индивидуальными привычками: насекомые ведь тоже способны к обучению.
Экспериментально показано, что видообразованию у цветковых растений могут способствовать такие изменения внешнего вида или аромата цветов, которые ведут к смене насекомого-опылителя (см. Необычный аромат гибридных орхидей способствует видообразованию, «Элементы», 11.06.2010; Чтобы избежать межвидовой гибридизации, орхидеям достаточно по-разному пахнуть, «Элементы», 15.06.2011). Хопкинс и Раушер не только продемонстрировали механизм усиления в действии, но и показали, что изменение цветка может способствовать видообразованию, даже если насекомые-опылители остаются прежними. Работа проливает дополнительный свет на причины удивительного разнообразия цветковых растений, развившегося во многом благодаря коэволюции с насекомыми-опылителями.
В отличие от «гена яркости», роль «гена оттенка» осталась невыясненной. Оттенок не влияет на выживаемость и плодовитость. Бабочки его игнорируют, поэтому вероятность гибридизации тоже не зависит от оттенка — только от яркости окраски. Почему «красный» аллель h распространился в симпатрической зоне, неясно. В настоящее время он, по-видимому, бесполезен для флоксов. Возможно, он был полезен в прошлом. Например, какой-то вид опылителей, ныне вымерший или ставший редким в этом районе, мог различать и запоминать оттенки и летать с красных цветов на красные или с синих на синие.
Источник: Robin Hopkins, Mark D. Rausher. Pollinator-Mediated Selection on Flower Color Allele Drives Reinforcement // Science. 2012. V. 335. P. 1090–1092.
См. также:
Генетическая несовместимость нарастает по параболе, «Элементы», 26.09.2010.
О том, как встреча после разлуки завершает видообразование, популярный синопсис Елены Бадьевой по статье В. А. Лухтанова «Правило Добржанского и видообразование путем усиления презиготической репродуктивной изоляции в зоне вторичного контакта популяций», «Журнал общей биологии», том 71, 2010. № 5, Сентябрь–октябрь. Стр. 372–385.
Новые виды образуются очень быстро, если меняется система распознавания половых партнеров, «Элементы», 30.04.2010.
-
1. А какой биологический смысл самим бабочкам игнорировать часть своей пищевой базы? (яркие, или наоборот, бледные цветы флоксов)?
2. Или цветовые предпочтения бабочек жестко заданы и не могут изменяться под давлением естественного отбора (доступной пищевой базы)?
3. Или бабочки находятся в условиях изобилия ресурсов и поэтому могут привередничать?
4. Или имеется в виду, что яркая окраска возникла (у одного из видов флоксов) лишь очень недавно, и бабочки еще не успели перестроиться?
5. Правда ли, что лишь в совпадающей части ареала один из флоксов имеет яркие цветы? Или яркие цветы бывают и в других популяциях флоксов?
6. А если не бывают, то, собственно, почему? Что удерживает дикие цветы флокса в одинаковой цветовой гамме на всем ареале? Ведь, во-первых, в искусственных условиях этот вид флокса демонстрирует весьма впечатляющую изменчивость окраски венчика, а во-вторых, сами исследователи установили, что нет никакой разницы в выживаемости и успешности размножения, например, между растениями со светло-красными или светло-синими цветками.
Так демонстрирует ли природа (в случае флоксов) ожидаемые микроэволюционные процессы по окраске венчика?
7. Почему в статье нет никаких статистических данных, демонстрирующих описываемую разницу в поведении опылителей? (ведь это здесь самое главное)-
-
1. бабочки не "игнорируют" часть своей пищевой базы, а скорее следуют простому правилу: "если я сейчас пила нектар из цветка с такой-то окраской, и мне понравилось, и вокруг много разных цветков и есть из чего выбрать, то лучше выбрать такой же цветок, как в прошлый раз". Это простейший и широко распространенный в животном мире алгоритм принятия решений: если какое-то действие было успешным, то лучше всего в следующий раз в аналогичной ситуации просто повторить это действие, а не искать приключений. Разумеется, у бабочек нет строгих предпочтений, и они не отказываются от нектара из других цветков. Но описанного алгоритма достаточно, чтобы обеспечить преимущество ярким цветкам в ситуации, описанной в статье.
2. Перелеты опылителей подробно описаны в дополнительных материалах, которые находятся в открытом доступе. Можно было бы и посмотреть самому, прежде чем выражать все эти сомнения.
3. В аллопатрической части ареала иметь нестандартную окраску цветам невыгодно, потому что при такой психологии опылителей они будут посещать необычный цветок реже других. И переносить пыльцу чаще всего с одного необычного на другой необычный, что чревато близкородственными скрещиваниями. Поэтому отбор в аллопатрической части должен способствовать конформизму окраски.
4. Для других читателей должен пояснить, что г-н Imperor - личность в узких кругах известная, и цель его комментариев - вовсе не поиск научной истины. Вступать с ним в дальнейшие дискуссии я не буду.-
Статью пока не читал, но новость интересная! Спасибо.
И, собственно, почему решил написать комментарий. "г-н Imperor", на мой взгляд, весьма интересный человек. То, что его взгляды кардинально расходятся с Вашими - еще не повод писать, что "цель его комментариев - вовсе не поиск научной истины". Если он действительно злостный тролль - докажите это. Давление на читателей и отсечение таким образом неугодных собеседников - не слишком красивая стратегия.-
"докажите это"
По-вашему, выкладывать здесь компромат на этого господина - более красивая стратегия? Но раз вы так настаиваете, могу сказать, например, что он помогал креационисту А. Милюкову писать большой и весьма яростный текст с "опровержениями" эволюции, за что Милюков выражал ему благодарность. Для меня достаточно этого факта, чтобы не общаться. Он не только в биологии, но даже и в квантовой физике (будучи биологом!) пытается "опровергнуть основы" и доказать "несостоятельность господствующих взглядов". Только поэтому Ваши с ним взгляды на половой отбор и совпали, кстати.-
1. Здесь Александр немного перепутал. Соответствующий "весьма яростный текст" Милюкова (насколько я понял) был написан, в основном, не с целью "опровержения эволюции", а с целью подвергнуть "весьма яростной критике" тот уровень "доказательств эволюции" (к сожалению, во многих местах действительно откровенно слабый), который Александр продемонстрировал в своих "Доказательствах эволюции".
Кстати, Александр, раз уж речь зашла об этом. Вы знаете, что у бделлоидных коловраток (проблему размножения которых Вы "успешно закрыли" лихим "кавалерийским маневром" с высыханием...) что в этом таксоне имеются и виды, НЕ имеющие ангидробиоза?
Причем эти виды процветают - они чрезвычайно распространены в большинстве водоемов земного шара. Вы об этом знали, когда выдвигали соответствующий тезис в качестве "доказательств верности теории"?
Я уж не говорю про Ваши удивительные рассуждения насчет "бессознательного отбора, производимого людьми" VS "настоящей селекции, производимой муравьями". Я не говорю про Ваших павлиних, которые (опять) отбирали там у Вас хвосты павлинам... вопреки результатам соответствующих исследований (см. пост ниже) и т.д.
Зачем было нужно приводить "доказательства", которые являются заранее провальными, я не знаю, поскольку боюсь предположить, что это было сделано с сознательной целью ввести читателей в заблуждение.
2. По поводу "квантовой физики". Здесь опять ошибка. Г-н Imperor вовсе не "опровергает основы", а просто является сторонником следующих физических концепций: 1) петлевой квантовой гравитации, 2) цифровой физики. Смысловое содержание этих двух концепций легко можно найти в сети. Таким образом, единственное преступление, которое совершил в данной области г-н Imperor - это то, что он пришел к личному выводу о справедливости этих концепций по дилетантски, т.е. путем достаточно простых рассуждений в стиле натурфилософии.
-
-
-
-
-


Александр, просто поразительно, настолько хорошо Вы знаете, каким именно образом рассуждают бабочки. Мне остаётся лишь позавидовать Вам в этом отношении.
Однако вот эта Ваша мысль уже менее удачна:
…Это простейший и широко распространенный в животном мире алгоритм принятия решений: если какое-то действие было успешным, то лучше всего в следующий раз в аналогичной ситуации просто повторить это действие, а не искать приключений...
Здесь я рекомендую Вам почитать соответствующий учебник. Например, «Экология. Особи, популяции и сообщества» (надеюсь, авторов не надо приводить?).
Раздел «Поведение хищников» и в частности, параграф 9.3.1.
Здесь Вы узнаете о многих полезных вещах. Например, о том, что помимо времени самого поглощения пищи (животным), имеется еще и время, которое животное тратит на переход (перелет, переползание) от одного пищевого объекта к другому. В это время (во время перехода) животное НЕ получает энергию.
Поэтому (с позиций голой биологической целесообразности) животному выгодно максимально сокращать длительность (и расстояние) такого перехода. Я надеюсь, всё это понятно?
Теперь, допустим, на искомом лугу имеется примерно равное число ярких и неярких цветов примерно равной пищевой ценности, которые распределены на этом лугу примерно равномерно. Представили?
Тогда бабочка, предпочитающая питаться ТОЛЬКО неяркими (или только яркими) цветами, потратит на переходы между своими пищевыми объектами примерно в ДВА раза больше времени, чем бабочка, садящаяся на оба типа цветов.
И только в том случае, если на лугу имеется ИЗОБИЛИЕ пищи, т.е. соседние цветы в изобилии находятся на расстоянии прямой и чёткой видимости (бабочки), и при этом высока вероятность, что эти соседние цветы еще не посещались конкурентами - только в этом случае наша привередливая бабочка сможет проложить себе подходящий маршрут, проходящий только по цветам, чья цветовая гамма нравится лично ей, не прибегая к дополнительным затратам на перелеты.
Но даже и в этом случае, дополнительные затраты всё равно будут иметь место. Это затраты на психические усилия по распознаванию и выделению именно определенных цветов (и определенного маршрута) из общей совокупности, где на самом деле ВСЕ цветы вполне подходящие!
Очевидно, что с точки зрения биологической целесообразности, такое поведение бабочки является слегка биологически НЕцелесообразным.
А если цветов на лугу меньше и значительная часть из них находится вне зоны прямой и чёткой видимости и(или) имеется много конкурирующих потребителей нектара - то такая привередливость бабочки будет уже крайне биологически нецелесообразной.
Я надеюсь, всё это достаточно ясно?
Т.е. возникает вопрос - а зачем, собственно, бабочка действует по определенному алгоритму (предложенному выше А.Марковым в виде "бабочкиных дум"), если в данной ситуации можно действовать вообще без всякого алгоритма (просто пересаживаясь на ближайшие цветы)? Ведь избирательные действия в данном конкретном случае не имеют смысла, приводя лишь к необоснованным дополнительным затратам (у этой особо привередливой бабочки). Неужели естественный отбор не способен отменить (или заблокировать) алгоритм избирательного поведения, бессмысленный в данном конкретном случае?
Вот поэтому в соответствующем параграфе уже озвученного мной учебника и написано следующее:
«…Другими словами, хищники должны специализироваться тогда, когда выгодные типы пищи многочисленны и (или) велики различия в выгодности, и жертвы должны поедаться безвыборочно тогда, когда выгодные категории редки и (или) различия в выгодности незначительны.»
Вот это пример БОЛЕЕ строгих рассуждений в рамках биологической целесообразности. Ваше же личное объяснение этого довольно удивительного (с точки зрения биологической целесообразности) пищевого поведения бабочек - представляет собой классический пример поверхностных интерпретационных схем, которыми так печально знаменита теория естественного отбора – когда те или иные факты из живой природы (нередко даже противоположные) легко «объясняются» (задним числом) разными интерпретационными схемами, которые удобно «вытащить из рукава» в том или ином конкретном случае.
"...бабочки не "игнорируют" часть своей пищевой базы, а скорее следуют простому правилу:
если я сейчас пила нектар из цветка с такой-то окраской, и мне понравилось, и вокруг много разных цветков и есть из чего выбрать, то лучше выбрать такой же цветок, как в прошлый раз"
Вот так. Написал (биолог!) несколько строк и успокоился...
И неважно, что возникает ощущение, будто наша бабочка:
1) существует в одном экземпляре,
2) в одном поколении,
3) присутствует на пятнадцатиминутной презентации
На самом же деле предполагаются:
1) целые популяции разных бабочек,
2) индивидуальная изменчивость бабочек по степени их пищевой "привередливости",
3) как минимум, прошедшие века, т.е. сотни сменившихся поколений бабочек,
4) естественный отбор тех бабочек, чьи пищевые предпочтения оказались самыми рациональными в данных условиях.
И почему, собственно, не написать было (А. Маркову) немного другие "бабочкины думы"?
Например, вот такие:
"..если я сейчас пила нектар из цветка с такой-то окраской (неяркой) И ЕЩЕ попила нектар из цветка с другой окраской (яркой)... и мне понравилось в ОБОИХ случаях, то лучше выбрать такие же цветки, как в прошлый раз - то есть И яркие И неяркие".
Вот это и огорчает в теоретических построениях любителей "всеобщей биологической целесообразности" - какие хотим "размышления бабочек" (какие нам удобны в данном конкретном случае) - такие размышления мы им и приписываем. Причем (часто) в разных случаях - прямо противоположные вещи пишем.
И снова хороший пример тех самых интерпретационных схем (легковесных), которые вытаскиваются из рукава, чтобы объяснить какую-то группу фактов. При этом одновременно "в другом рукаве" имеется другая интерпретационная схема, которая позволяет объяснить группу уже других фактов, нередко вообще противоположных.
Вот и здесь, Александр, Вы сейчас рассказываете нам, что на всем протяжении своего ареала (!) который в случае обсуждаемого нами вида флокса простирается чуть ли не на половину Северной Америки - окраска венчика этого вида везде одинакова потому, что это растение таким образом борется: 1) за внимание опылителей (которые в свою очередь теоретически страдают от недостатка пищи!), 2) против близкородственного скрещивания!
И именно поэтому (по Вашему мнению) предполагаемые микроэволюционные процессы (по окраске венчика) в разных популяциях флоксов пресекаются "в зародыше" на всём протяжении ареала вида...
Но при этом в ДРУГИХ своих текстах Вы с удовольствием рассуждаете о симпатрическом видообразовании, о сотнях (!) видов цихлид в африканских озерах... и никакая "угроза близкородственного скрещивания" им почему-то (в этом видообразовании) не грозит...
И одновременно (с третьей стороны) мы знаем об удивительном видовом разнообразии и разнообразии окрасок цветов у покрытосеменных растений. В том числе, и у предельно близких в таксономическом отношении, например, у обсуждающихся здесь же (!) близких видов флоксов. Интересно, как же такое разделение могло произойти, если оно (по А. Маркову) должно было пресекаться на самых ранних стадиях "психологией опылителя"? (термин А. Маркова)
И, наконец, одновременно (с четвертой стороны) все мы знаем о существовании (и биологическом процветании) многих видов самоопыляющихся растений.
Александр, я не знаю, что там у Вас за «узкие круги» такие. Остаётся только надеяться, что эти «круги» достаточно узки :)
А вот по поводу «поиска научной истины» - это Вы явно перегибаете палку.
Тут уж, скорее, дело обстоит с точностью до наоборот.
К сожалению, я вынужден наблюдать, как Вы (начиная с некоторого времени) слишком увлеклись легковесным редукционизмом, которым злоупотребляет теория естественного отбора. Увлеклись настолько, что в угоду этой теории стали нередко грешить той самой «научной истиной» за чистоту которой Вы столь демонстративно боретесь.
Например, я с чрезвычайным удивлением прочитал (в Вашем же блоге) Ваши рассуждения о том, что вполне достаточно на словах набросать какую-нибудь логическую схему «в рамках биологической целесообразности»… и ничего эмпирически проверять и доказывать уже не надо.
Объясните мне, почему Вы продолжаете рассказывать читателям о павлинихах, которые отобрали павлинам их хвосты, если эти рассуждения НЕ соответствуют результатам исследований (причем не только в отношении павлинов, но и почти во всем таксоне куриных)?
Где же Ваше стремление к научной истине?
Я понимаю, что рассказывать о результатах, которые противоречат принятой Вами концепции – не слишком приятно, но хотя бы упоминать о наличии подобных противоречащих фактов Вы ведь должны (в стремлении к научной истине)?
Почему Вы упорно продолжаете рассказывать читателям о том, что привлекательные черты людей – это свидетельство «хороших генов», когда такие заявления, во-первых, НЕ подтверждаются результатами соответствующих исследований, а во-вторых, имеются исследования (в том числе, и наши), что эстетические предпочтения вообще не имеют отношения к каким бы то ни было «хорошим генам», а являются, скорее всего, следствием неких глубинных психических эффектов, вообще не имеющих никакого отношения к биологической целесообразности?
Вы можете быть не согласны с такой точкой зрения. Действительно, можно сказать, что вопрос до конца еще не решен. Но пока ведь все накопленные факты говорят против Ваших рассуждений, и одновременно ничего не говорит «ЗА» (кроме чисто теоретических рассуждений в рамках концепции естественного отбора). Так почему же Вы об этом даже не упоминаете?
Вы с удовольствием рассказываете читателям о том, что «только самые приспособленные самцы шалашников» (с самыми "хорошими генами") могут откладывать красные цветочки к красным цветочкам, а синие ракушки – к синим ракушкам…
И при этом, во-первых, не чувствуете, насколько смехотворно всё это звучит, а во-вторых, просто не замечаете вопросов (и не отвечаете на них), которые тут же возникают, если мы начинаем рассуждать в подобном стиле:
http://elementy.ru/news?chapter=20370&discuss=431742
К сожалению, я так и не нашел (видимо, плохо умею). Не могли бы Вы (из уважения к читателям) дать конкретную ссылку на эти дополнительные материалы (ведь в тексте Вашей заметки нет ни соответствующих данных, ни соответствующих ссылок).
Последние новости










Одомашненные представители вида Phlox drummondii. Изображение с сайта www.thompson-morgan.com