Микротомография ископаемых мшанок
Анна Коромыслова, Зоя Толоконникова, Петр Федоров, Алексей Пахневич
«Природа» №1, 2021
 Анна Викторовна Коромыслова — кандидат геолого-минералогических наук, старший научный сотрудник Палеонтологического института имени А. А. Борисяка РАН. Область научных интересов — морфология, систематика, палеогеография мшанок ордовика и мела. |
 Зоя Алексеевна Толоконникова — кандидат геолого-минералогических наук, доцент кафедры региональной и морской геологии Кубанского государственного университета. Специалист в области изучения мшанок позднего палеозоя (морфология, систематика, стратиграфическое значение). |
 Петр Владимирович Федоров — кандидат геолого-минералогических наук, доцент кафедры осадочной геологии Института наук о Земле Санкт-Петербургского государственного университета. Научные интересы связаны с вопросами стратиграфии, палеонтологии, седиментологии. |
 Алексей Валентинович Пахневич — кандидат биологических наук, старший научный сотрудник Палеонтологического института имени А. А. Борисяка РАН и Объединенного института ядерных исследований. Область научных интересов — девонские, каменноугольные и современные брахиоподы, рентгеновская томография, кристаллографическая текстура биогенного минерального вещества. |
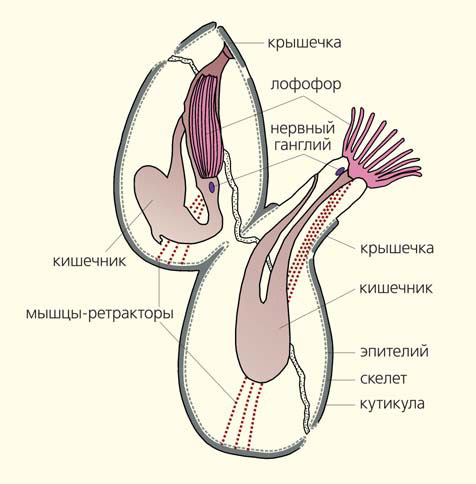
Мшанки (тип Bryozoa) — это прикрепленные колониальные бентосные беспозвоночные (преимущественно морские, реже пресноводные), питающиеся органическими частицами, которые они вылавливают из водной взвеси при помощи венчика щупалец — лофофора. Эти животные появились на Земле в начале ордовика (около 487 млн лет назад), они были разнообразно представлены в водных бассейнах прошлого и живут сегодня. Насчитывается более 6 тыс. видов современных мшанок и более 15 тыс. ископаемых видов [1]. Среди мшанок выделяют три класса: филактолемата (Phylactolaemata), стенолемата (Stenolaemata) и гимнолемата (Gymnolaemata). Филактолематы — это пресноводные мшанки. У представителей этого класса, включающего около 100 современных видов, скелет образован мягкой кутикулой, поэтому их колонии не сохраняются в ископаемом состоянии. Однако статобласты (спящие почки) филактолемат, покрытые хитиновой оболочкой, известны с пермского периода [2]. Стенолематы — это морские мшанки, обладающие кальцитовым скелетом. Они доминировали в палеозое (с раннего ордовика до перми) и продолжают существовать в современных морях [3, 4]. В палеозое гимнолематы были представлены сверлящими мшанками отряда ктеностомата (Ctenostomata). Отряд сохранился и ныне. Ктеностоматы — это морские, реже пресноводные мшанки, не имеющие минерального скелета. В поздней юре (мезозой) появилась другая группа гимнолемат — отряд хейлостомата (Cheilostomata), которая начиная с позднего мела и поныне является доминирующей и морфологически самой разнообразной группой морских мшанок. Представители хейлостомат могут иметь кальцитовый, арагонитовый или биминеральный скелет [5].
Колонии бывают размером от первых миллиметров до нескольких сантиметров. Но некоторые достигают значительных размеров, как, например, арктическая Tabulipora с толщиной ветвей до 7 см [1]. Продолжительность жизни мшанок одних современных видов менее 6 мес, тогда как другие могут жить 50 лет [1, 6].
Колонии состоят из многочисленных крошечных особей размером менее 1 мм (их называют зооидами), которые выполняют различные функции. Основу колоний составляют автозооиды — самостоятельно питающиеся и размножающиеся особи. В колонии также присутствуют один или несколько типов гетерозооидов, отличающиеся от автозооидов как строением, так и выполняемой функцией. На протяжении фанерозоя в колониях мшанок были развиты гетерозооиды многих различных типов, из которых в данной статье будут упомянуты только неозооиды, эксилязооиды и авикулярии. Неозооиды и эксилязооиды известны только у палеозойских стенолемат. Предполагают, что они служили для обеспечения прочности колонии, а также для увеличения расстояния между лофофорами автозооидов (возможно, это позволяло оптимизировать питание всей колонии). Авикулярии появились в колониях хейлостомат в начале мелового периода, и они многообразны у современных представителей этого отряда. Авикулярии, аналогично различным шипам (в том числе акантостилям палеозойских стенолемат), защищают колонии мшанок как от питающихся ими животных (например, кольчатых червей и рыб), так и тех, которые просто засоряют колонии (например, личинок донных животных, оседающих на мшанках).
В колониях в среднем насчитывается несколько десятков или сотен тысяч зооидов, но возможны и исключения. Например, колония Flustra foliacea весом всего 13 г образована около 1 333 000 зооидами [6], тогда как мшанка Monobryozoon состоит всего из одного зооида [1, 3].
Как изучают ископаемые мшанки?
Исследование скелетов мшанок разных геологических эпох имеет свои отличительные особенности. Колонии постпалеозойских (мезозойских и кайнозойских) мшанок в основном изучают при помощи сканирующего электронного микроскопа, поскольку систематика многих групп этих мшанок базируется преимущественно на внешних морфологических признаках. В основу традиционной систематики палеозойских мшанок положены представления о внутриколониальных структурах. Их описания составляют по ориентированным тонким срезам (шлифам), которые изучают с помощью светового микроскопа. Недостаток подобного метода — разрушение ископаемой колонии. Кроме того, перекристаллизация вещества скелета зачастую может приводить к утрате систематически важных структур и признаков. Достижения технического прогресса неизбежно приводят к появлению новых методов в исследовании окаменелостей. Один из таких новых методов — это рентгеновская компьютерная микротомография. Мы полагаем, что в будущем этот метод исследования будет приобретать все большее значение для исследования палеонтологических объектов. Метод позволяет выявлять разницу в минеральном составе и плотности окаменелости и вмещающей породы.
Ж. Роджер и Э. Бьюдж [8] первыми использовали рентгеновские снимки для изучения кайнозойских мшанок Cellepora, обраставших раковины брюхоногих моллюсков. Получены уже первые результаты использования микротомографии для изучения ордовикских [9, 10, 11], каменноугольных [12, 13], пермских [10], юрских [14], меловых [15–19] и современных [20 и др.] мшанок. Как оказалось, применение этого метода значительно расширило возможности ученых. Микротомография позволяет получить тысячи виртуальных срезов колонии в различных плоскостях, которые могут заменять шлифы. На основе виртуальных срезов создаются и трехмерные изображения самих колоний, которые по своей информативности превосходят изображения, полученные с помощью сканирующего микроскопа. К тому же при микротомографии колонии не разрушаются, что особенно важно при изучении музейного материала.
В эволюционном развитии мшанок было несколько переломных рубежей, не всегда совпадающих с коренными перестройками в биоте Земли: ранний и начало среднего ордовика — этап ранней эволюции мшанок; поздний девон — пермь — расцвет палеозойских мшанок, достижение их максимального разнообразия; поздний мел — формирование морфологического разнообразия мезокайнозойских мшанок. Для исследования мшанок мы применяем комплексный подход, сочетающий сканирующую микроскопию и микротомографию с изучением прозрачных ориентированных шлифов в оптическом микроскопе. В статье мы продемонстрируем результаты, полученные методом микротомографии при изучении ископаемых мшанок, в частности представителей палеозойских стенолемат и позднемеловых хейлостомат.
Колонии мшанок, изображенные в настоящей статье, сканировали с помощью рентгеновского компьютерного микротомографа Skyscan 1172 (Bruker Corporation) в Центре рентгеноструктурных дифракционных исследований Ресурсного парка Санкт-Петербургского государственного университета (Санкт-Петербург) и в Палеонтологическом институте имени А. А. Борисяка РАН (Москва).
Древнейшие мшанки
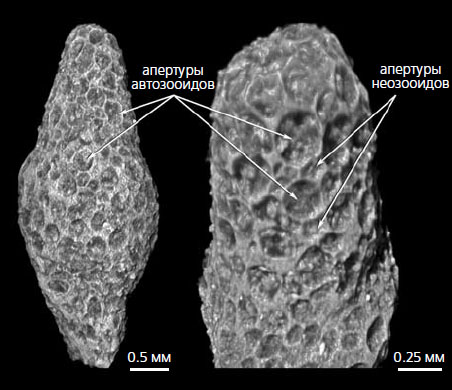
Изображения древнейшей мшанки Revalotrypa inopinata, полученные с помощью метода рентгеновской компьютерной микротомографии; нижний ордовик, Ленинградская обл. [9]
Предполагается, что мшанки могли существовать уже в кембрии, но тогда они еще не имели минерального скелета [21]. Самые древние достоверные находки мшанок имеют возраст около 485 млн лет и происходят из тремадокского яруса нижнего ордовика Южного Китая. Они представлены несколькими видами родов Prophyllodictya, Nekhorosheviella, Orbiramus [22]. Мшанки возрастом 477 млн лет были обнаружены нами [9] в глауконитовых песчаниках флоского яруса нижнего ордовика на территории Ленинградской обл., которая тогда располагалась в центре палеоконтинента Балтика. Древнейшие мшанки Балтики, Revalotrypa inopinata и R. yugaensis, имели очень мелкие колонии (не более 5 мм в высоту) и состояли из автозооидов и неозооидов.
Представители рода Planopora возрастом около 470 млн лет — одни из древнейших билатеральных мшанок. Они были обнаружены в одном из карбонатно-глиняных иловых холмов («геккеровых горбов»), формировавшихся одновременно с накоплением вмещающих слоистых глауконитовых известняков дапинского яруса среднего ордовика на территории Ленинградской обл. [10, 11]. В их плоских колониях отверстия зооидов (их называют апертурами) открывались на обе стороны колонии. Апертуры автозооидов Planopora имеют лунарии (серповидные структуры, влияющие на ориентацию щупалец лофофора). Разделены автозооиды пузырчатой тканью, которая служила для укрепления колонии и увеличения расстояния между автозооидами. В отличие от мшанок Revalotrypa gibbosa, полусферические колонии которых часто крепились к раковинам брахиопод, мшанки Planopora volkhovensis, а также булавовидные Esthoniopora clara имеют в своем основании холдфаст — прикрепительную структуру, охватывающую цилиндрический объект или несколько разноориентированных объектов, что, вероятно, свидетельствует о возможном их креплении к спикулам губок. В отложениях «геккерова горба» найдены также уникальные колонии Hemiphragma insolitum, обраставшие фосфатные трубки широко распространенного палеозойского морского животного сфеноталлус (Sphenothallus), который, возможно, принадлежал к книдариям [23].
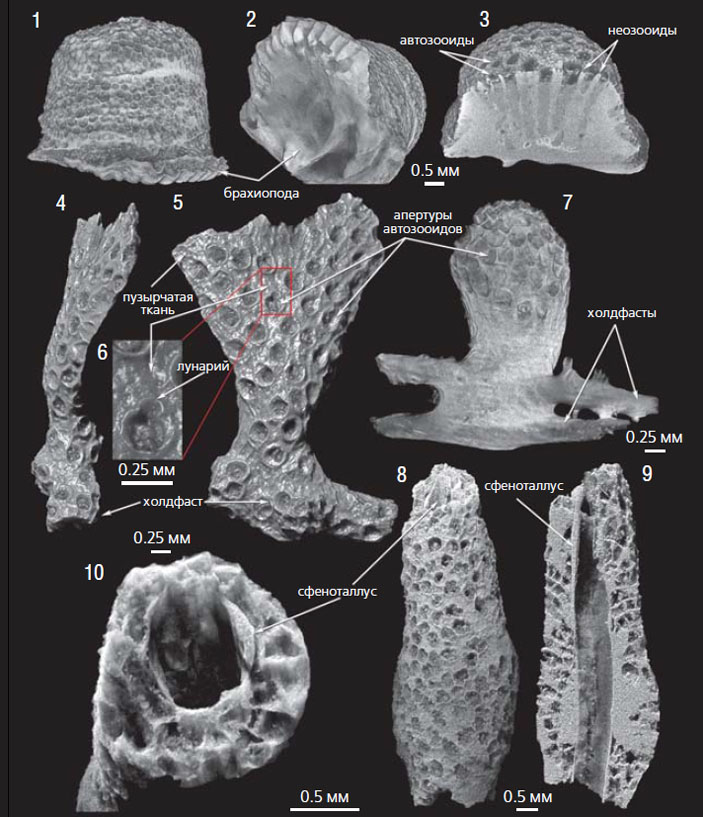
Изображения дапинских мшанок из «геккерова горба», полученные методом микротомографии; средний ордовик, Ленинградская обл. [10, 11]: 1–3 — Revalotrypa gibbosa, обраставшая раковину брахиоподы; 4–6 — Planopora volkhovensis и 7 — Esthoniopora clara, крепившиеся к спикулам губок; 8–10 — Hemiphragma insolitum, обраставшая трубку сфеноталлуса
Перечисленные выше находки ордовикских мшанок пока крайне редки, некоторые из них представлены единичными экземплярами. Метод микротомографии позволил получить многочисленные виртуальные срезы колоний в разных плоскостях, без их разрушения, а также построить 3D-модели самих колоний.
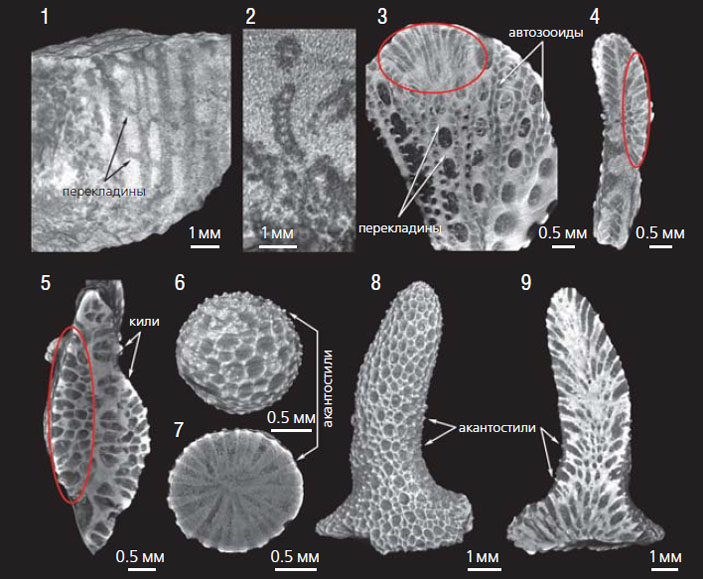
Изображения мшанок из нижнего карбона Южного Казахстана (1, 2) и средней перми Самарской обл. России (3–9), полученные методом микротомографии [10, 13]: 1 — сетчатая колония Rectifenestella cf. longacella; 2 — сетчатая колония Narynella cf. narynica; 3–5 — срезы воронкообразной колонии Wjatkella wjatkensis с инкрустирующей колонией Dyscritella incrustata (выделено красным овалом); 6, 7 — сферическая колония D. incrustata; 8, 9 — ветвистая колония D. spinigeriformis
Мшанки позднего палеозоя
Изменение внешнего облика колоний за счет появления новых внутренних структур в позднем палеозое могло быть результатом комплексного воздействия биотических и абиотических факторов. Максимума разнообразия мшанки достигли к началу каменноугольного периода (358,9 млн лет назад). Наивысший расцвет характерен для мшанок отряда фенестрата (Fenestrata), особенно для родов Rectifenestella, Spinofenestella и Fenestella, которые образовывали различные сетчатые колонии: вееровидные, пластинчатые, ветвистые, перистые, воронковидные и др. Форма камер автозооидов первостепенна для определения систематического положения мшанок этого отряда. Внешне похожие сетчатые колонии, относящиеся к неродственным таксонам, обладают разными внутренними структурами, что демонстрирует важность использования метода микротомографии для изучения этих мшанок.
Мы изучили мшанки космополитного рода Rectifenestella из Южнокаратауского палеобассейна [13]. На снимках видны сетчатые колонии, образованные бифуркирующими прямыми прутьями с двумя рядами автозооидов пятиугольного сечения и узкими косыми перекладинами без зооидов. Из того же палеобассейна была изучена Narynella cf. narynica с одним рядом округлых или трапециевидных в среднем сечении автозооидов на прутьях сетки. Благодаря соединению слабоволнистых прутьев узкими перекладинами через неравномерные промежутки образовывалась крупнопетлистая колония.

Изображения позднемеловых мшанок, полученные методами сканирующей электронной микроскопии (1, 2, 5, 6, 9, 10) и микротомографии (3, 4, 7, 8, 11, 12) [15–17]: 1–4 — Pachydermopora grodnoensis из кампана Белоруссии: 1 — общий вид колонии, 2 — два зооида, 3 — продольный срез зооидов, 4 — 3D-модель зооида; 5–8 — Beisselinopsis quincunx из маастрихта Луганской обл. Украины: 5 — общий вид колонии, 6 — зооид, 7 — продольный срез зооидов, 8 — 3D-модель зооида; 9–12 — Acoscinopleura occulta из маастрихта Германии: 9 — общий вид колонии, 10 — зооид, 11 — продольный срез зооидов, 12 — 3D-модель зооида
Фенестраты господствовали в бентосных сообществах шельфа и материкового склона до конца пермского периода, зачастую успешно конкурируя не только с мшанками, относящимися к иным отрядам, но и с другими группами организмов (брахиоподами, моллюсками, иглокожими). К середине перми (270,6 млн лет назад) колонии фенестрат стали более массивными за счет увеличения количества рядов автозооидов на прутьях. У некоторых родов развивались суперструктуры, образование которых связано с разрастанием килей на прутьях, разделяющих ряды автозооидов. Кили, срастаясь вместе, формировали защитную сетку над сетчатой колонией. На большей части Восточно-Европейского палеобассейна в казанском веке обитала мшанка Wjatkella wjatkensis [24, 25]. Для нее были типичны воронкообразные колонии с прутьями, несущими 2–4 ряда четырех-шестиугольных в срединном сечении автозооидов, и с широкими короткими перекладинами между ними. Исследование образцов колоний этих мшанок из казанских отложений бассейна р. Сок (Самарская обл.) с помощью микротомографии показало обилие крупных бугорков на высоких килях [10]. Кили и бугорки, кроме выполнения защитной функции, вероятно, были важны для регуляции гидродинамической ситуации вокруг автозооидов и усиления эффективности питания колонии [26].
В казанском веке среднепермской эпохи в Восточно-Европейском палеобассейне кроме фенестрат существовали и представители отряда трепостомата (Trepostomata), постепенно теряющие свою доминантную позицию в мшанковых сообществах с конца девонского периода. На восточной окраине палеобассейна численно преобладали трепостоматы из рода Dyscritella [24, 25]. Примечательно, что за 218 млн лет существования дискрителл (с девона до конца триаса) произошла смена более чем 120 видов этого рода, причем наибольшее разнообразие видов пришлось на пермский период (71% видового богатства). Для этих мшанок характерны ветвистые, массивные и инкрустирующие колонии. Форма колоний зависела частично от субстрата, на котором изначально закреплялась анцеструла (первый зооид, дающий начало всей колонии). Так, мшанка Dyscritella incrustata, обнаруженная в казанских отложениях бассейна р. Сок, возможно, еще при жизни обрастала сетку Wjatkella wjatkensis. Благодаря применению микротомографии удалось проследить последовательное зарастание автозооидами D. incrustata петель колонии W. wjatkensis. Кроме того, на изображениях колонии D. incrustata, полученных с помощью микротомографии, различимы два типа акантостилей (они имеют разные размеры), выступающих над поверхностью колонии. Их обилие и хаотичное расположение можно рассматривать либо как способ пассивной защиты от хищников, либо как приспособление для снижения давления воды на колонию [27]. У Dyscritella spinigeriformis из той же сокской мшанковой ассоциации форма колонии ветвистая, но при этом акантостили также дифференцированы. На микротомограмме видов Dyscritella обнаружены обильные эксилязооиды в межзооидальных промежутках.
Несмотря на разнообразие адаптивных возможностей у палеозойских мшанок, к концу пермского периода большинство стенолемат навсегда исчезли из морских бассейнов планеты. Конец палеозойской эры стал переломным — произошла смена доминантов в мшанковых ассоциациях на уровне групп и зарождение мезозойской фауны.
Мшанки позднего мела
Ближе к середине позднего мела (86,3 млн лет назад) в морях стали доминировать мшанки отряда Cheilostomata, многие из которых имели сложноустроенные колонии. С помощью сканирующей микроскопии и микротомографии мы изучали хейлостомат из отложений кампанского и маастрихтского ярусов Евразии — строение их мелких колоний (менее 15 мм в высоту) в целом, а также морфологию отдельных их зооидов [15–18].
У некоторых хейлостомат, например Beisselina, Pachydermopora и Beisselinopsis, фронтальная поверхность автозооидов сильно обызвествлена, поэтому строение колонии скрыто. С помощью микротомографии была изучена форма камер автозооидов, а также пространственное расположение перистома (трубка для выхода лофофора), выводковых камер и других структур, имеющих систематическое значение для этих мшанок [15, 17].
Особенность хейлостомных мшанок рода Acoscinopleura — присутствие одной или нескольких каверн в криптоцисте (известковой пластине, которая подстилала фронтальную мембрану зооидов). Подобные каверны не встречаются у современных мшанок, поэтому их функция неясна. Обычно каверны в колониях Acoscinopleura видны на поверхности криптоцисты, так как ее тонкий слой, закрывавший каверну, часто не сохраняется в ископаемом состоянии. Однако у мшанок A. occulta каверны в криптоцисте были обнаружены только при микротомографии [16]. Это связано с тем, что часть криптоцисты, закрывающая каверны снаружи, оказалась толще, чем у многих представителей этого рода, и не разрушилась в процессе фоссилизации.
Исследование мшанок Tobolocella levinae с помощью метода микротомографии позволило установить в их колониях, помимо автозооидов и авикуляриев, новый тип зооидов (он был назван Т-зооидом) [18]. Т-зооиды и авикулярии образуют ряды, которые чередуются с рядами автозооидов. Т-зооиды меньше автозооидов и не имеют выводковых камер. Они могли быть разновидностью автозооидов (если мягкое тело, заполнявшее их, было таким же, как и в автозооидах) и также выполнять функцию питания колонии. Но, возможно, их мягкое тело было изменено, и по аналогии с авикуляриями они выполняли функцию защиты колонии. Также они могли увеличивать расстояние между лофофорами автозооидов и тем самым улучшать питание колонии.

Колонии мшанок Tobolocella levinae из маастрихта Северного Казахстана [18]: 1 — общий вид колонии, 2 — зооид, изображения получены с помощью сканирующей микроскопии; 3 — продольный срез зооидов, 4 — 3D-модель зооидов, изображения получены с помощью микротомографии
Минеральный состав скелета меловых мшанок
Одно из самых распространенных химических соединений, из которых беспозвоночные строят минеральный скелет, — это карбонат кальция. Обычно он представлен двумя минералами — кальцитом и арагонитом, сходными по химическому составу, но различающимися строением кристаллической решетки. Кальцит чаще, чем арагонит, встречается в составе скелета беспозвоночных. Феномен объясняется меньшей устойчивостью арагонита: этот минерал может легко разрушаться или замещаться другими минералами после смерти организма. Однако есть животные с биминеральным скелетом, где присутствуют сразу оба минерала. Таковы, например, многие двустворчатые моллюски, в том числе хорошо известные мидии. Перламутр мидий образован арагонитом, а внешний минеральный слой — кальцитом.
Как мы уже упоминали, среди мшанок минеральным скелетом обладают все представители стенолемат и хейлостоматы. Предполагается, что биоминерализация скелета в этих группах мшанок возникала независимо друг от друга, у стенолемат в раннем ордовике, а у хейлостомат — в поздней юре. Все стенолематы имеют кальцитовый скелет, тогда как у хейлостомат он может быть кальцитовым, арагонитовым или биминеральным. Предполагается, что биминеральные виды хейлостомат появились еще в маастрихтском веке позднего мела [5]. Исключение — колонии ордовикских стенолемат, в стенках зооидов которых обнаружен фосфат кальция [28]. Помимо световых микроскопов минеральный скелет мшанок изучают с помощью методов рентгеновской порошковой дифракции, лазерной рамановской спектроскопии, сканирующей электронной микроскопии (в том числе с полевой эмиссией), микрозондового анализа, дифракции обратно рассеянных электронов [5], реже компьютерной рентгеновской микротомографии [29].
Интересные факты зачастую появляются случайно. Так произошло и с коллекцией мшанок рода Acoscinopleura из Зенкенбергского музея (Франкфурт-на-Майне, Германия), которая была изучена с помощью микротомографии [16]. На томографических срезах некоторых колоний A. crassa (из отложений маастрихтского яруса Германии) было выявлено много контрастных участков, неравномерно распределенных по скелету колонии. Большая часть колонии на срезах была темно-серой, а по контуру внутренней полости зооидов и вокруг латеральных и дистальных септул (структур, обеспечивающих связь зооидов) наблюдался контрастный светло-серый слой, толщина которого варьирует от 0,02 до 1,14 мм. Несмотря на то что контрастные участки хорошо видны на изображениях [16], эта особенность не была отмечена при описании A. crassa.

Скелет мшанки Acoscinopleura crassa, маастрихт, Хеммор, Германия [16]: 1 — общий вид колонии, снятый с помощью сканирующего микроскопа; 2–7 — результаты микротомографии: 2 — общий вид колонии, прозрачность 90%, 3–5 — продольные срезы вдоль узкой стороны колонии, 6 — продольный срез вдоль широкой стороны, 7 — поперечный срез колонии
Возможны две интерпретации появления контрастного слоя. Первый вариант — контурный (при вращении объекта на 180°) или круговой (при вращении объекта на 360°) артефакт, который выглядит как тонкий контрастный слой на краю образца. Такие артефакты иногда образуются в процессе микротомографического исследования образца на границе раздела образец / воздух. Причина их возникновения — нарушение настроек оборудования. Однако мы предполагаем, что в нашем случае контрастный слой в скелете мшанок не артефакт. Вращение образца производилось на 180°, поэтому круговой артефакт вокруг округлых структур, в данном случае септул, образоваться не мог. Контрастный слой толще обычного контурного артефакта и распределен неравномерно. Его толщина варьирует, особенно сильно вокруг септул. Есть участки колонии, где этот слой не просматривается. В целом из 110 экземпляров хейлостомных мшанок, которые изучались на микротомографе в Палеонтологическом институте РАН, только у двух экземпляров A. crassa обнаружилась такая особенность.
Второй вариант интерпретации появления контрастного слоя — биминеральный скелет мшанок A. crassa (данный факт ранее не указывался). Увеличение контрастности может быть связано с появлением в стенках зооидов колонии примеси или другого минерала, который контрастен с кальцитом или арагонитом. При использованных параметрах томографии (сила тока — 100 мА, напряжение — 100 кВ) эти минералы между собой неконтрастны. Но либо один из них мог быть замещен другим минералом, содержащим химические элементы с большим атомным номером, либо в кристаллической решетке первоначального минерала могли накопиться ионы-примеси подобных элементов, увеличивающих контрастность. Хорошо известно, что в карбонатах кальций иногда замещается ионами Mg, Sr, Ba, Pb, Zn, Co, Na, U, Fe, Mn, Cu [30].
Остается открытым вопрос, когда произошло замещение контрастным минералом или накопление примесей — в прижизненном состоянии или уже после смерти животного в ходе диагенетических изменений осадочной породы. Возможен и тот и другой вариант. Важно, что контрастность наблюдается не во всей колонии, а только на некоторых участках стенок зооидов. Это значит, что скелет исходно состоял из минералов, имеющих разные кристаллические решетки, т.е. результаты описанных наблюдений можно рассматривать как косвенное доказательство биминеральности скелетов A. crassa. Для уточнения вещественного состава скелетов этих мшанок нужен, безусловно, дальнейший их химический и минеральный анализ.
Ранее хейлостомные мшанки с биминеральным скелетом, жившие в низких широтах, были описаны только из отложений палеогена. Тем не менее предполагалось, что появление биминеральных скелетов должно было произойти ранее — еще в маастрихте, т.е. еще в мезозое [5]. Наши данные подтверждают это предположение, но описанные выше мшанки обитали в высоких широтах. В более ранних публикациях [31] указывалось, что только внешний слой фронтальной поверхности зооидов хейлостомных мшанок мог состоять из арагонита. Однако у изученных мшанок A. crassa контрастный слой в основном оконтуривает полость зооидов, реже присутствует во внешнем слое их фронтальной поверхности. Таким образом, можно предположить, что у A. crassa арагонит мог присутствовать во внутренних слоях скелета.
***
Мшанки — интересная и перспективная группа беспозвоночных, позволяющая проводить эволюционные, морфологические и другие исследования в силу своего высокого разнообразия, адаптивности и длительного существования. Использование комплекса разнообразных методов, в том числе сканирующей электронной микроскопии и рентгеновской компьютерной микротомографии в сочетании с классической оптической микроскопией, повышает ценность результатов их исследования для биологов и палеонтологов.
Работа поддержана Российским фондом фундаментальных исследований (проект 18-05-00245-А). Часть исследований выполнена в рамках государственной программы повышения конкурентоспособности Казанского (Приволжского) федерального университета среди ведущих мировых научно-образовательных центров.
Литература
1. Gordon D. P., Taylor P. D., Bigey F. P. Phylum Bryozoa. New Zealand inventory of biodiversity. 1. Animalia. Radiata, Lophotrochozoa, Deuterostomia. D. P. Gordon (ed.). Canterbury, 2009: 271–297.
2. Виноградов А. В. Новые ископаемые мшанки континентальных водоемов азиатской части России и Казахстана // Палеонтологический журнал. 1995; 4: 43–53.
3. Вискова Л. А. Морские постпалеозойские мшанки. М., 1992. (Труды Палеонтологического института РАН. Т. 250).
4. Горюнова Р. В. Филогения палеозойских мшанок. М., 1996. (Труды Палеонтологического института РАН. 267).
5. Taylor P. D., James N. P., Bone Y. et al. Evolving mineralogy of cheilostome bryozoans // Palaios. 2009; 24: 440–452.
6. Клюге Г. А. Мшанки северных морей СССР. М.; Л., 1962.
7. Астрова Г. Г., Шишова Н. А. Наставление по сбору и изучению ископаемых мшанок. М., 1963.
8. Roger J., Buge É. L’association cellepore-gastropode dans les faluns de la Touraine // Bulletin de la Société Géologique de France Série. 1947; 5(17): 461–470.
9. Fedorov P. V., Koromyslova A. V., Martha S. O. The oldest bryozoans of Baltoscandia from the lowermost Floian (lower Ordovician) of north-western Russia: two new rare, small and simple species of Revalotrypidae // Paläontologische Zeitschrift. 2017; 91: 353–373.
10. Коромыслова А. В., Федоров П. В., Толоконникова З. А. Результаты исследования ордовикских и пермских мшанок методом рентгеновской компьютерной микротомографии // Труды Палеонтологического общества. Т. 3. М., 2020; 23–31.
11. Koromyslova A. V., Fedorov P. V. The oldest bifoliate cystoporate and two other bryozoan taxa from the Dapingian (Middle Ordovician) of north-western Russia // Journal of Paleontology. 2021; 95(1): 24–39. DOI: 10.1017/jpa.2020.73.
12. Wyse Jackson P. N., McKinney F. K. A micro-computed tomography and scanning electron microscopy investigation of the structure of Polyfenestella Bancroft, 1986 (Bryozoa: Fenestrata), from the Mississippian of Scotland: revealing the nature of its heteromorphs // Irish Journal of Earth Sciences. 2013; 31: 19–24.
13. Tolokonnikova Z., Fedorov P., Wyse Jackson P. N. Bryozoan fauna from the Mississippian of the Akuyuk section, southern Kazakhstan // Neues Jahrbuch für Geologie und Paläontologie — Abhandlungen. 2020; 295(2): 159–168.
14. Вискова Л. А., Пахневич А. В. Новая сверлящая мшанка из средней юры Московской области и ее микротомографическое исследование // Палеонтологический журнал. 2010; 2: 38–46.
15. Коромыслова А. В., Пахневич А. В. Новые виды Pachydermopora Gordon, 2002 и Beisselina Canu, 1913 (Bryozoa: Cheilostomida) из кампанского отторженца Белоруссии и их микротомографическое исследование // Палеонтологический журнал. 2016; 1: 40–50.
16. Koromyslova A. V., Martha S. O., Pakhnevich A. V. The internal morphology of Acoscinopleura Voigt, 1956 (Cheilostomata, Bryozoa) from the Campanian and Maastrichtian of Central and Eastern Europe // Paläontologische Zeitschrift. 2018; 92: 241–266.
17. Koromyslova A. V., Martha S. O., Pakhnevich A. V. Revision of the Porina-like cheilostome Bryozoa from the Campanian and Maastrichtian of Central Asia // Annales de Paléontologie. 2019; 105: 1–19.
18. Koromyslova A. V., Pakhnevich A. V., Fedorov P. V. Tobolocella levinae n. gen., n. sp., a cheilostome bryozoan from the late Maastrichtian of northern Kazakhstan: scanning electron microscope and micro-CT study // Neues Jahrbuch für Geologie und Paläontologie — Abhandlungen. 2019; 294(1): 91–101.
19. Martha S. O., Ruthensteiner B., Taylor P. D., Hillmer G., Matsuyama K. Description of a new cyclostome species from the middle Santonian of Germany using micro-computed tomography // Australasian Palaeontological Memoirs. 2019; 91–99.
20. Matsuyama K., Titschack J., Baum D., Freiwald A. Two new species of erect Bryozoa (Gymnolaemata: Cheilostomata) and the application of non-destructive imaging methods for quantitative taxonomy // Zootaxa. 2015; 4020: 81–100.
21. Taylor P. D., Waeschenbach A. Phylogeny and diversification of Bryozoans // Palaeontology. 2015; 58(4): 585–599.
22. Ma J.-Y., Taylor P. D., Xia F., Zhan R. The oldest known bryozoan: Prophyllodictya (Cryptostomata) from the lower Tremadocian (Lower Ordovician) of Liujiachang, south-western Hubei, central China // Palaeontology. 2015; 58: 925–934.
23. Vinn O., Kirsimäe K. Alleged cnidarian Sphenothallus in the Late Ordovician of Baltica, its mineral composition and microstructure // Acta Palaeontologica Polonica. 2015; 60: 1001–1008.
24. Морозова И. П. Мшанки поздней перми. М, 1970. (Труды Палеонтологического института АН СССР. Т. 122).
25. Tolokonnikova Z. Permian bryozoans from the Nemda horizon (Roadian) of Samara Region, Russia // Paläontologische Zeitschrift. 2020; 94(1): 79–92.
26. Suárez Andrés J. L., Wyse Jackson P. N. Feeding currents: a limiting factor for disparity of Palaeozoic fenestrate bryozoans // Palaeogeography, Palaeoclimatology, Palaeoecology. 2015; 433: 219–232.
27. Ernst A. Bryozoa of the Upper Permian Zechstein Formation of Germany // Senckenbergiana lethaea. 2001; 81(1): 135–181.
28. Conti S., Serpagli E. Bimineralic (calcareous and phosphatic) skeleton in Late Ordovician Bryozoa from Sardinia: geological implication // Bollettino della Societa Paleontologica Italiana. 1988; 27(2): 129–162.
29. Jacob D. E., Ruthensteiner B., Trimby P. et al. Architecture of Anoteropora latirostris (Bryozoa, Cheilostomata) and implications for their biomineralization. Scientific Reports. 2019; 9(11439).
30. Мак-Коли Дж. Кальцита группа // Минералогическая энциклопедия. Л., 1985; 95–99.
31. Benedix G., Jacob D. E., Taylor P. D. Bimineralic bryozoan skeletons: a comparison of three modern genera // Facies. 2014; 60: 389–403.













Схема организации зооидов мшанок — с втянутым лофофором (в верхней части) и с вытянутым и развернутым лофофором [7, с изменениями]