Титанозавры России
Александр Аверьянов
«Природа» №10, 2019

 Об автореАлександр Олегович Аверьянов — профессор РАН, доктор биологических наук, заведующий лабораторией териологии Зоологического института РАН, профессор Института наук о Земле Санкт-Петербургского государственного университета. Научные интересы связаны с морфологией и эволюцией позвоночных животных. |
Завроподы — группа нептичьих* динозавров, включающая крупных и очень крупных растительноядных животных. Некоторые завроподы достигали 30 м в длину и 80 т веса и были самыми крупными наземными животными за всю историю Земли. Эти ящеры ходили на четырех колонноподобных ногах. Для них были характерны маленькая голова и очень длинная шея (у некоторых видов она была в четыре-пять раз длиннее туловища). В настоящее время известно уже около 200 родов завропод, что составляет примерно 15–20% разнообразия всех нептичьих динозавров. Однако, несмотря на обилие находок, завроподы долгое время были наименее изученной группой динозавров. В частности, филогенетические связи разных их групп стали проясняться лишь в последнее время благодаря внедрению кладистического метода.

Основные клады завропод [15, рис. 14]
Самую раннюю радиацию завропод представляют архаичные формы позднего триаса — ранней юры (вроде вулканодона из ранней юры Зимбабве). Они отличались от своих предков — «прозавропод» — более крупными размерами и хождением на четырех колонноподобных ногах. Однако эти первые архаичные завроподы еще плохо изучены. В частности, неизвестно строение их черепа. Большинство «настоящих» завропод относятся к кладе Eusauropoda, радиация которой началась в ранней юре. В качестве примеров первых эузавропод приведу шунозавра из ранней юры Китая, барапазавра из ранней юры Индии, ферганозавра из средней юры Киргизии [1] и патагазавра из средней юры Аргентины. Характерные признаки эузавропод это, в частности, не менее 13 шейных позвонков, укороченные и редуцированные фаланги конечностей и более совершенная зубная система (при обработке пищи зубы верхней и нижней челюстей тесно соприкасались и образовывали фасетки стирания). Для всех представителей этой клады характерна морщинистая скульптура эмали зубов, что позволяет легко диагностировать их изолированные зубы. К первым эузавроподам относятся также маменьсизавриды (Mamenchisauridae) с необычной даже для завропод длинной шеей, включавшей 15 и более позвонков. Некоторые виды маменьсизавра из поздней юры — раннего мела Китая могли достигать 30 м в длину и более 70 тонн веса. Маменьсизавриды были распространены преимущественно в Китае и Юго-Восточной Азии. Лишь недавно ископаемые остатки представителя этой группы нашли в среднеюрских отложениях Западной Сибири [2]. Это самые древние находки завропод с территории нашей страны.
Более эволюционно продвинутые завроподы относятся к кладе Neosauropoda, которая разделяется на две большие клады — Diplodocoidea и Macronaria. Неозавроподы появились в средней юре, они характеризуются, в частности, отсутствием зубчиков по краям зубных коронок и формированием полукруглого вертикального блока из пястных костей, в результате чего передняя конечность опиралась только на фаланги пальцев, а не на всю ладонь. Общие для диплодокоидов признаки — это тонкие зубы-колышки и длинный хвост, кончик которого похож на хлыст. К диплодокоидам относятся три семейства (реббахизавриды, дикраеозавриды и диплодоциды), радиация которых происходила в поздней юре — раннем мелу в Северной и Южной Америке и в Африке.
Другую большую кладу неозавропод составляют макронарии, названные так по большим носовым отверстиям. Камаразавр из поздней юры США — наиболее примитивный представитель макронариев. Брахиозавриды и более продвинутые макронарии объединяют в кладу Titanosauriformes, широко распространенную в поздней юре — мелу на всех континентах, кроме Антарктиды. К этой кладе относятся, например, брахиозавр и жираффатитан (брахиозавриды) из поздней юры соответственно США и Танзании, эухелоп (эухелопиды) из раннего мела Китая и эркету из самого начала позднего мела Монголии. Большинство диплодокоидей и нетитанозавровых макронарий вымерло к началу позднего мела.

Титанозавр рюкватитан из мела Танзании. Реконструкция А. А. Атучина
Для позднего мела характерна радиация последней большой группы завропод — титанозавров. Позднемеловые титанозавры известны из Южной Америки, Африки, Мадагаскара, Индии, Европы, Центральной и Восточной Азии. В Северной Америке завроподы вымерли в конце раннего мела, и лишь один род (аламозавр) в самом конце позднего мела проник на этот континент из Южной Америки.
Титанозавры — последние гиганты
Титанозавры — единственная группа завропод, дожившая до конца мелового периода (маастрихта), причем в то время они были распространены почти на всех континентах и были достаточно разнообразны. Маастрихтские титанозавры известны в Северной (аламозавр) и Южной Америке (гондванатитан), на Мадагаскаре (рапетозавр), в Индии (изизавр), в Европе (ампелозавр, магиарозавр) и в Азии (нэмэгтозавр, опистоцеликаудия). К титанозаврам относится почти половина всех известных родов завропод. Неудивительно, что при таком разнообразии и размеры представителей этой группы варьируют больше, чем в других группах завропод. Среди них есть как сравнительно небольшие животные (сальтазавр, неукуэнзавр), так и самые крупные завроподы (аргентинозавр, пуэртозавр и др.).
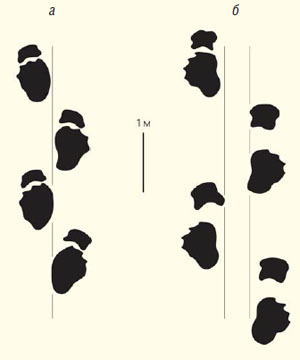
Следовые дорожки нетитанозаврового завропода (а) и титанозавра (б) [16, рис. 1]
Титанозавры характеризуются рядом морфологических особенностей, которые отличают их от других завропод. Напомню, что для динозавров характерна парасагиттальная постановка конечностей, при которой конечности ставятся близко к средней линии тела. Такая же постановка наблюдается и у териевых млекопитающих, тогда как у ящериц, крокодилов и однопроходных млекопитающих конечности расставлены широко. Особенности постановки конечностей хорошо видны на следовых дорожках, оставленных динозаврами. У нетитанозавровых завропод отпечатки передней и задней конечности расположены близко к средней линии следовой дорожки, часто ее пересекают. У титанозавров конечности были расставлены заметно шире, и их следы расположены на существенно большем расстоянии от средней линии следовой дорожки. Более широкая постановка конечностей обеспечивалась, в частности, ориентировкой бедренной кости не вертикально, а под углом к вертикальной оси тела, что позволяло вынести голень дальше от средней линии тела. Благодаря такой постановке бедра у титанозавров образовывался особый латеральный вырост на бедренной кости, служивший местом крепления мышц, а поперечное сечение самой кости было эллипсовидным, вытянутым в медиолатеральном направлении, что увеличивало ее прочность. С широкой постановкой конечностей также связана серия морфологических модификаций в передней конечности и в грудной клетке. Видимо, с особенностями локомоции титанозавров связано укорочение их хвоста, который состоял примерно из 35 позвонков. Передние хвостовые позвонки у всех титанозавров (кроме опистоцеликаудии) процельные, т.е. вогнутые спереди и выпуклые сзади. У представителей клады продвинутых титанозавров-литостротий (Lithostrotia) процельными были не только передние, но и средние хвостовые позвонки (форма же сочленения задних позвонков могла сильно варьировать). Литостротии также характеризуются наличием небольших костных бляшек в коже — остеодерм. Опистоцеликаудия уникальна тем, что у нее передние хвостовые позвонки опистоцельные (т.е. выпуклые спереди и вогнутые сзади), а средние хвостовые — платицельные (плоские спереди и сзади). Укорочение хвоста и его увеличенная подвижность за счет появления шаровидного сустава между позвонками, возможно, были связаны с функцией хвоста как дополнительной опоры тела, что позволяло титанозаврам приподнимать переднюю часть тела и передние конечности во время питания или спаривания. Также у титанозавров передний отросток подвздошной кости таза ориентирован не вертикально, как у других завропод, а горизонтально, выступая наружу. Еще одна уникальная особенность титанозавров — неокостенение элементов запястья, кисти и предплюсны, которые, видимо, оставались хрящевыми в течение всей жизни.

Бедренная кость диплодока (а) и сальтазавра (б). У титанозавра сальтазавра бедренная кость расположена под углом α к вертикальной оси тела и имеется выступ на боковой поверхности, обозначенный стрелкой [17, рис. 4]
Функциональное значение многих особенностей скелета титанозавров пока полностью не понятно. Возможно, расширение постановки конечностей и латеральный изгиб подвздошной кости были связаны с увеличением туловища титанозавров, в частности с чрезмерным развитием органов пищеварения (желудок, кишечник), которые могли использоваться как бродильные чаны для переваривания зеленых частей растений с помощью микроорганизмов. Как бы то ни было, эти морфологические особенности обеспечили титанозаврам определенный эволюционный успех и позволили им пережить другие группы завропод.
Находки титанозавров в России
Долгое время завроподы в России были известны лишь по очень редким и фрагментарным остаткам, не поддающимся определению. Зубы завропод из раннемелового местонахождения Тээтэ в Якутии были отнесены к американскому роду камаразавр [3] (однако изолированные зубы завропод не могут быть диагностированы на родовом уровне, так что данную находку можно в действительности определить не ближе, чем Macronaria indet). Это местонахождение завропод — самое северное в Северном полушарии. Известно оно также уникальной находкой зубов детенышей завропод — первым свидетельством того, что эти динозавры могли размножаться в приполярных широтах.
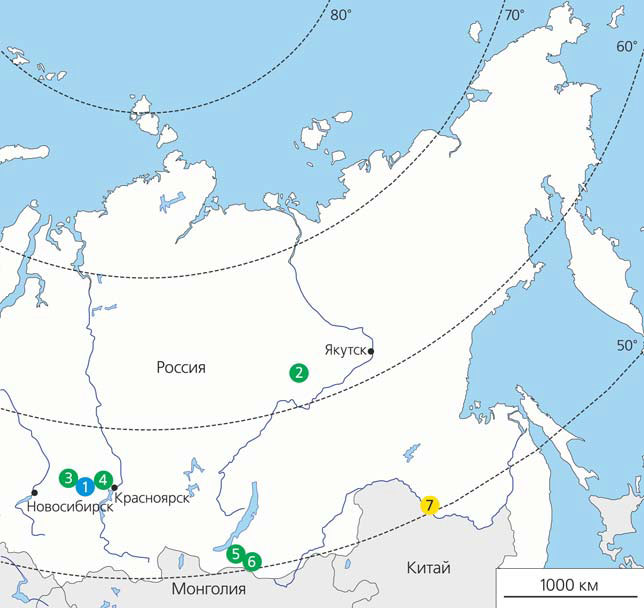
Места находок завропод в Сибири: 1 — Березовский карьер (итатская свита, батский ярус средней юры), 2 — Тээтэ (батылыхская свита, берриасский — барремский ярус раннего мела), 3 — Шестаково 1 и 3 (илекская свита, аптский ярус раннего мела), 4 — Большой Кемчуг 3 (илекская свита, барремский? ярус раннего мела), 5 — Могойто (муртойская свита, барремский и аптский? ярусы раннего мела), 6 — Красный Яр (хилокская свита, аптский ярус раннего мела), 7 — Благовещенск (удурчуканская свита, маастрихтский ярус позднего мела)
Другая интересная находка ископаемых остатков завропод в России — серия хвостовых позвонков из нижнемеловых отложений Сланцевого рудника близ г. Ульяновска, на берегу Волги, найденных еще в 1982 г. В. М. Ефимовым. Уникальность ее в том, что она найдена в морских отложениях с богатой фауной беспозвоночных, а это позволяет точно датировать находку верхним готеривом (зона Speetoniceros versicolor). Большинство других находок завропод сделано в континентальных отложениях, возраст которых часто определяется в довольно широких пределах. Морфология позвонков также оказалась уникальной, что позволило выделить эту форму в новый род и вид Volgatitan simbirskiensis [4]. Сильно выраженная процельность передних и средних хвостовых позвонков свидетельствует о принадлежности волгатитана к титанозаврам клады Lithostrotia. Однако, в отличие от многих титанозавров, на нижней стороне позвонков у волгатитана имеется продольный гребень, а не борозда. Также для волгатитана характерны дополнительные сочленения на невральной дуге передних хвостовых позвонков, которые ранее были обнаружены только у одного представителя титанозавров — у эпахтозавра из верхнего мела Аргентины. Проведенный филогенетический анализ позволил поместить волгатитана в основание филогенетической линии, на вершине которой находятся гигантские титанозавры из позднего мела Южной Америки, достигавшие 60–70 т веса (аргентинозавр, нотоколосс, патаготитан, пуэртазавр). Однако по расчетам масса самого волгатитана составляла всего около 17 т. Очевидно, что эта филогенетическая линия титанозавров в раннем мелу была широко распространена, но в позднем мелу сохранилась только в Южной Америке, где в условиях начавшейся изоляции эти титанозавры приобрели гигантские размеры. Волгатитан — древнейший известный в мире титанозавр.

Передний хвостовой позвонок волгатитана: вид снизу (а), сверху (б), сзади (в), сбоку (г) и спереди (д)
Разрозненные зубы и хвостовые позвонки завропод находили также в раннемеловом местонахождении Могойто близ о. Гусиного в Бурятии [5, 6]. Недавно по хвостовым позвонкам из Могойто был описан новый титанозавр Tengrisaurus starkovi [7]. У тенгризавра и передние, и средние хвостовые позвонки глубоко процельные, что свидетельствует о его принадлежности к группе литостротий. Невральная дуга передних хвостовых позвонков тенгризавра сильно пневматизирована, т.е. снабжена серией особых полостей с отверстиями вдоль остистого отростка. Это эволюционно продвинутый морфологический признак, который встречается еще только у некоторых титанозавров из позднего мела Южной Америки. Существование столь продвинутого титанозавра в раннем мелу Азии подчеркивает неполноту наших знаний о ранних этапах эволюции титанозавров.

Средний хвостовой позвонок тенгризавра: вид спереди (а), сверху (б), сбоку (в), сзади (г) и снизу (д)
Эволюционно продвинутые титанозавры характеризуются тонкими карандашеподобными зубами с круглой или эллиптической апикальной фасеткой стирания. Такие зубы очень похожи на зубы диплодокоидов, но они, безусловно, появились у титанозавров независимо. Для более примитивных титанозавров характерны зубы с более широкими коронками и V-образными боковыми фасетками стирания. У промежуточных по эволюционному положению титанозавров зубы тонкие, но встречаются оба варианта фасеток [8]. Интересно, что зубы у титанозавров, найденные в Могойто, были примитивного типа: коронка у них весьма широкая, хорошо выражены боковые фасетки стирания V-образной формы. Эти зубы могли принадлежать тенгризавру, описанному по хвостовым позвонкам из данного местонахождения. Но если это так, то наблюдается своеобразный эволюционный диссонанс между примитивными зубами и продвинутыми хвостовыми позвонками. Однако в одной экосистеме могли сосуществовать два таксона завропод и даже более. Единственный зуб завропода из местонахождения Красный Яр в Бурятии принадлежит более продвинутому титанозавру: коронка зуба еще достаточно толстая, но имеет одну апикальную фасетку стирания эллиптической формы. Эта фасетка расположена под тупым углом к вертикальной оси зуба, тогда как у более продвинутых титанозавров фасетка стирания почти перпендикулярна продольной оси зуба.

Зубы титанозавров из местонахождений Могойто (а–в) и Красный яр (г–е). Каждый зуб изображен с трех сторон [7, 18]
Одно из крупнейших местонахождений динозавров в нашей стране — это Шестаковский комплекс местонахождений близ дер. Шестаково в Кемеровской обл. Наиболее значимые местонахождения комплекса — Шестаково 1 (обрыв вдоль р. Кия) и Шестаково 3 (искусственная дорожная выемка к югу от дер. Шестаково). Это одно из немногих мест, где встречаются скелеты динозавров в сочленении. В частности, в Шестаково 3 известно уникальное захоронение скелетов сибирского пситтакозавра [9, 10]. Шестаковские местонахождения относятся ко второй половине раннего мела, когда по всей Центральной и Восточной Азии были широко распространены фауны пситтакозаврового комплекса. Ископаемые остатки других динозавров на Шестаковских местонахождениях довольно редки. Тем не менее, в ходе многолетних работ сотрудники Томского государственного университета нашли в Шестаково 1 почти полную стопу завропода [11]. На берегу р. Кия (Шестаково 1) практически каждый год вымываются крупные кости завропод, в частности, найдены шейные и грудные позвонки, а также почти полный крестец. По этим остаткам недавно был описан новый таксон завропод — Sibirotitan astrosacralis [12]. Филогенетический анализ позволил определить сибиротитана как нетитанозаврового титанозавриформа (т.е. принадлежащего к кладе Titanosauriformes, но не к Titanosauria). Для сибиротитана характерно наличие всего пяти крестцовых позвонков, тогда как у большинства титанозавриформ их шесть. Другая его отличительная черта — схождение крестцовых ребер в одну точку. Такая особенность отмечалась ранее только у эпахтозавра и эухелопа. С эухелопом сибиротитана также роднят некоторые особенности строения шейных позвонков. Так что, видимо, филогенетическое положение сибиротитана было определено правильно. Однако для сибиротитана неизвестно строение хвостовых позвонков, столь важное в систематике титанозавров.
Долгое время в местонаходении Шестаково 3 не встречалось никаких ископаемых остатков завропод. Лишь в прошлом году сотрудники Кемеровского краеведческого музея нашли здесь первые их кости — хвостовые позвонки и шевроны (гемальные дуги). Передние и средние хвостовые позвонки глубоко процельные, они однозначно принадлежат титанозавру из клады литостротий, т.е. таксону более эволюционно продвинутому, чем сибиротитан из соседнего местонахождения. Шевроны имеют уникальную морфологию, известную ранее только для диплодокоидов и не встреченную ни у одного представителя титанозавров. Если предположить, что данные позвонки и шевроны принадлежат сибиротитану, то тогда его филогенетическое положение было определено неверно. Однако в действительности между местонахождениями Шестаково 1 и 3 нет общих форм позвоночных (кроме пситтакозавра), и оба местонахождения могут иметь разный геологический возраст. В таком случае существование особого таксона завропод в Шестаково 3 более вероятно. Кроме того, как уже отмечалось, в былых экосистемах вполне могли сосуществовать два или более таксона завропод. Решить загадку помогут только новые находки. Процельный хвостовой позвонок литостротиевого титанозавра известен также из раннемелового местонахождения Большой Кемчуг 3 в Красноярском крае.
Завроподы позднемелового возраста на территории России пока практически неизвестны. Единственная находка — фрагмент карандашеподобного зуба, принадлежавший эволюционно продвинутому титанозавру из местонахождения Благовещенск, которое датируется самым концом мелового периода — маастрихтским веком [5]. Завроподы России находятся пока на самой начальной фазе изучения. Но уже первые находки позволяют существенно скорректировать представления об эволюции титанозавров. Так, например, раньше безоговорочно принималась гипотеза о южноамериканском происхождении титанозавров [13]. Теперь же, в связи с находками древнейших титанозавров в России, можно уверенно говорить об азиатском происхождении группы. Безусловно, систематические работы на старых местонахождениях и открытие новых местонахождений позволят прояснить эволюцию этих гигантов на шестой части суши.
Работа выполнена при финансовой поддержке Российского научного фонда (грант 19-14-00020).
Литература
1. Alifanov V. R., Averianov A. O. Ferganasaurus verzilini, gen. et sp. nov., a new neosauropod (Dinosauria, Saurischia, Sauropoda) from the Middle Jurassic of Fergana Valley, Kirghizia // Journal of Vertebrate Paleontology. 2003; 23(2): 358–372.
2. Averianov A. O., Krasnolutskii S. A., Ivantsov S. V. et al. Sauropod remains from the Middle Jurassic Itat Formation of West Siberia, Russia // PalZ. 2019. DOI: 10.1007/s12542-018-00445-8.
3. Курзанов С. М., Ефимов М. Б., Губин Ю. М. Новые архозавры из юры Сибири и Монголии // Палеонтологический журнал. 2003; (1): 55–59.
4. Averianov A. O., Efimov V. M. The oldest titanosaurian sauropod of the Northern Hemisphere // Biological Communications. 2018; 63(3): 145–162. DOI: 10.21638/spbu03.2018.301.
5. Несов Л. А. Динозавры Северной Евразии: новые данные о составе комплексов, экологии и палеобиогеографии. СПб., 1995.
6. Averianov A. O., Starkov A. I., Skutschas P. P. Dinosaurs from the Early Cretaceous Murtoi Formation in Buryatia, Eastern Russia // Journal of Vertebrate Paleontology. 2003; 23(3): 586–594.
7. Averianov A. O., Skutschas P. P. A new lithostrotian titanosaur (Dinosauria, Sauropoda) from the Early Cretaceous of Transbaikalia, Russia // Biological Communications. 2017; 62(1): 6–18. DOI: 10.21638/11701/spbu03.2017.102.
8. Averianov A. O., Sues H.-D. Sauropod teeth from the Upper Cretaceous Bissekty Formation of Uzbekistan // Historical Biology. 2017; 29(5): 641–653. DOI: 10.1080/08912963.2016.1229777.
9. Лопатин А. В., Мащенко Е. Н., Тарасенко К. К. и др. Уникальное захоронение раннемеловых позвоночных в Западной Сибири (местонахождение Шестаково-3, Кемеровская область) // Доклады Академии наук. 2015; 462(5): 620–623.
10. Averianov A. O., Voronkevich A. V., Leshchinskiy S. V., Fayngertz A. V. A ceratopsian dinosaur Psittacosaurus sibiricus from the Early Cretaceous of West Siberia, Russia and its phylogenetic relationships // Journal of Systematic Palaeontology. 2006; 4(4): 359–395.
11. Averianov A. O., Voronkevich A. V., Maschenko E. N. et al. A sauropod foot from the Early Cretaceous of Western Siberia, Russia // Acta Palaeontologica Polonica. 2002; 47(1): 117–124.
12. Averianov A. O., Ivantsov S. V., Skutschas P. P. et al. A new sauropod dinosaur from the Lower Cretaceous Ilek Formation, Western Siberia, Russia // Geobios. 2018; 51(1): 1–14.
13. Gorscak E., O’Connor P. M. Time-calibrated models support congruency between Cretaceous continental rifting and titanosaurian evolutionary history // Biology Letters. 2016; 12(4). DOI: 10.1098/rsbl.2015.1047.
14. Averianov A. O., Skutschas P. P. Additions to the Early Cretaceous dinosaur fauna of Transbaikalia, eastern Russia // Proceedings of the Zoological Institute of the Russian Academy of Sciences. 2009; 313(4): 363–378.
15. Wilson J. A. Overview of sauropod phylogeny and evolution // The Sauropods Evolution and Paleobiology (Eds.: K. A. Curry Rogers, J. A. Wilson). Berkeley; Los Angeles; L., 2005; 15–49.
16. Wilson J. A., Carrano M. T. Titanosaurs and the origin of «wide-gauge» trackways: a biomechanical and systematic perspective on sauropod locomotion // Paleobiology. 1999; 25(2): 252–267. DOI: 10.1017/S0094837300026543.
* Нептичьими называют всех динозавров, кроме относящихся к группе птиц (Aves). С позиций филогенетической систематики птицы — одна из групп (клад) хищных динозавров (Theropoda).

























Реконструкция раннемелового ландшафта в районе местонахождения Шестаково, Кемеровская обл. Показаны, в частности, завропод сибиротитан и рогатый динозавр пситтакозавр сибирский. Рисунок А. А. Атучина