Инфекционность амилоидов
Александр Дергалёв
«Природа» №5, 2016
 Об автореАлександр Андреевич Дергалёв — аспирант и научный сотрудник Института биохимии им. А. Н. Баха в составе Федерального исследовательского центра «Фундаментальные основы биотехнологии» РАН. Область научных интересов — молекулярная генетика, амилоидные и полиглутаминовые белки, клеточная биология. Участник конкурса «Био/мол/текст» 2015 г.* |
Конформационные болезни человека — это нейродегенеративные патологии, связанные с изменением конформации некоторых белков, ее превращением из мономерной растворимой в полимерную фибриллярную. На протяжении многих лет такие заболевания было принято условно делить на два класса: прионные и амилоидные. Однако в последнее время появилось множество свидетельств того, что граница между ними несколько иллюзорна, а прионная концепция применима к большинству патологических амилоидов нервной системы человека. Изучая ненаследуемые амилоиды в дрожжах Saccharomyces cerevisiae, мы можем найти ответ на вопрос, существует ли принципиальная разница в строении инфекционных и неинфекционных амилоидных полимеров.
Начиная разговор о прионах и амилоидах, следует внести ясность в терминологию. Согласно современным традициям употребления этого термина, прионы — это самовоспроизводящиеся белковые изоформы, обладающие свойством инфекционности, т.е. способностью передаваться от клетки к клетке. На самом деле под это определение попадают белковые изоформы разной природы, однако объединяет их одно: все они проявляются как некие черты фенотипа (иногда губительные для клеток, иногда, наоборот, полезные), которые способны распространяться между клетками без участия наследственной информации, закодированной с помощью ДНК/РНК.

Амилоиды** — это белковые агрегаты с особым способом укладки молекул, при котором отдельные, амилоидогенные, участки образуют фибриллярные полимеры с кросс-β-структурой, т.е. состоящие из β-слоев, направленных перпендикулярно оси фибриллы (рис. 1) [1]. Отличить амилоидные полимеры от других форм белковой агрегации (например, аморфных агрегатов, образующихся при тепловом шоке) можно по нескольким признакам, помимо уже упомянутой структуры:
- амилоиды обладают повышенной прочностью и не растворяются в сильных ионных детергентах (таких, как додецилсульфат натрия) при комнатной температуре;
- амилоидные полимеры катализируют присоединение к себе свободных молекул того же белка и их глубокую структурную перестройку;
- амилоиды связываются со специфическим набором красителей, в частности с тиофлавином и конго красным (последний традиционно используют гистологи для обнаружения амилоидных отложений в тканях больного) (рис. 2);
- в отличие от образования актиновых филаментов и микротрубочек, полимеризация белка в амилоидную фибриллу происходит без участия молекул АТФ и ГТФ, поэтому такие фибриллы могут собираться (но не разбираться!) самопроизвольно, причем как в клетке, так и in vitro.

Рис. 2. Амилоидные отложения по периферии клеток жировой ткани, наблюдаемые с помощью светового (слева) микроскопа и поляризационного при окрашивании красителем конго красным. Размер одной клетки — около 100 мкм. Фото: pharmaceuticalintelligence.com
Способность тех или иных участков белка укладываться в амилоид зависит от их аминокислотного состава. Известны случаи, когда мутации в амилоидном домене белка существенно уменьшают его склонность переходить в фибриллярную форму [2].
Большинство известных на данный момент прионов имеют амилоидную природу: амилоидные фибриллы — тот самый инфекционный агент, который переносится от клетки к клетке и определяет их прионный фенотип. Однако известны и другие механизмы самокопирования белковых состояний, с амилоидностью никак не связанные. Например, дрожжевой прион β связан с активацией протеиназы Prb1. Она синтезируется неактивной благодаря блокирующему N-концевому пептиду. Без него, став активной, молекула Prb1 может активировать другие молекулы протеиназы, отщепляя у них этот пептид. Популяция молекул Prb1 может долго оставаться неактивной, но ее «инфицирование» лишь одной молекулой протеиназы без блокирующего пептида запускает цепную реакцию, активируя все молекулы этого белка.
Известные прионные заболевания, болезни Крейцфельдта — Якоба, куру, коровье бешенство и некоторые другие, вызваны одним и тем же инфекционным агентом — амилоидными полимерами белка PrP. Этот мембранный белок есть у всех млекопитающих, а также у птиц и у рыб, и в норме он выполняет в клетках не до конца изученные функции (которые иногда не очень грамотно приписывают прионной форме белка, в то время как выполняет их как раз таки неприонная форма). PrP при патологии сворачивается в амилоидную форму, которая вызывает гибель нейронов и может заражать другие клетки (а иногда и другие организмы). Интересно, что PrP способен укладываться в амилоид несколькими способами, образуя фибриллы с различными конформациями, а те, в свою очередь, ложатся в основу разных прионных штаммов, по-разному взаимодействуя с клеточным окружением амилоида. В конечном счете разница между конформациями амилоидов приводит к некоторым различиям в течении вызываемых ими прионных заболеваний.
Амилоиды нейродегенеративных заболеваний
Помимо болезней, традиционно рассматриваемых как прионные, в число конформационных заболеваний человека входит также множество других патологий нервной системы, развивающихся, как правило, с возрастом, — болезни Паркинсона, Альцгеймера, Хантингтона и др. Их развитие также подразумевает агрегацию собственных белков клетки (Aβ-пептидов, α-синуклеина, хантингтина и др.) в амилоидные фибриллы, которые, в свою очередь, собираются в губительные для клеток вне- и внутриклеточные амилоидные агрегаты. Известно даже, что промежуточная (на пути превращения из растворимого мономера в фибриллу) форма укладки Aβ-пептида, уже содержащая в своей структуре β-тяжи, тоже обладает токсическим действием на клетки [3].
Подобные болезни раньше не рассматривались с точки зрения прионной концепции: считалось, что такие амилоиды образуются в каждой нервной клетке автономно и не могут передаваться горизонтально, т.е. не обладают свойством инфекционности. Однако в последнее время накапливается множество свидетельств типично прионного характера распространения амилоидов внутри нервной ткани больных, а также перекрестного «затравления» полимеризации разных амилоидных белков человека — процесса, когда полимеры одного амилоидного белка, взаимодействуя с растворимой формой другого, запускают его агрегацию в фибриллы. Так, например, амилоидные агрегаты из Aβ-пептида могут «затравлять» полимеризацию тау-белка в нейрофибриллярные пучки [4]. Подобные наблюдения заставляют ученых взглянуть на болезни, всегда считавшиеся прионными, иначе. И сейчас в научной печати появляется все больше работ, в которых обсуждаются прионные (или прионоподобные) свойства амилоидов, вызывающих возрастные нейродегенеративные патологии.
Сейчас соотношение понятий «амилоид» и «прион» меняется (рис. 3). И хотя этот процесс иногда происходит больше в терминологической, нежели в экспериментальной, плоскости, особую важность приобретает вопрос: существуют ли какие-то принципиальные различия в молекулярной архитектуре фибрилл инфекционных и неинфекционных амилоидов? Существуют ли конформации, более других подходящие для горизонтального или вертикального распространения от клетки к клетке, и, наоборот, конформации, которые не в состоянии никого «заразить»? Как разные конформации амилоидов отличаются по восприимчивости к лекарствам? (Да, у некоторых прионных амилоидов, как и у бактерий, обнаружена способность развивать устойчивость к используемым против них лекарствам. Меняя свои свойства, эти амилоиды делают неэффективным лечение заболевания с помощью уже существующих средств и заставляют ученых изобретать новые — такая гонка вооружений в чем-то роднит прионы с другими видами патогенов.) В качестве модельного объекта для поиска ответов на эти вопросы хорошо подходят амилоиды дрожжей.

Рис. 3. Изменение соотношения понятий «амилоид» и «прион». В последние годы все большее число амилоидов обретает прионоподобный статус
Прионы и амилоиды дрожжей
Пекарские дрожжи Saccharomyces cerevisiae уже долгое время служат полезным инструментом не только в пищевой промышленности, но и в руках ученых — биохимиков, молекулярных биологов, генетиков. Высокая степень изученности дрожжей (их генов, белков, физиологических процессов), большая скорость роста, удобство культивирования — все это делает их удобным объектом для моделирования и изучения разнообразных явлений, от эндоцитоза до возникновения многоклеточности. Дрожжи — весьма популярный модельный объект и среди ученых, занимающихся конформационными болезнями [5]. Используя дрожжи, мы можем намного быстрее и проще исследовать молекулярные механизмы нейродегенерации — общие (или по крайней мере схожие) у дрожжей и человека.
Особое обстоятельство, расширяющее наши возможности в изучении амилоидов у дрожжей, — это наличие у них собственных прионов, большинство из которых имеет амилоидную природу. Таким образом, мы можем исследовать не только человеческие амилоиды и прионы, привнесенные в дрожжевую клетку (что, при всей несомненной пользе, все же немного искусственно), но и дрожжевые прионы в их естественной среде обитания. На данный момент известно больше десятка принципиально разных (вовлекающих различные белки) дрожжевых прионов, а некоторые из них к тому же имеют разные штаммовые варианты, т.е. амилоидные конформации одного и того же белка, по-разному себя проявляющие [6].
Важное отличие прионов дрожжей от прионов млекопитающих — куда более дружелюбное отношение к организму-хозяину. Если полимеризация белка PrP у млекопитающих приводит к развитию болезни и неотвратимой, пусть и не очень скорой, гибели носителя, то значительная часть дрожжевых прионов не особенно вредит клеткам (разве что несколько замедляет их рост), а в некоторых ситуациях даже выполняет в них жизненно важные функции. Другое отличие в том, что у прионов млекопитающих инфекционность заключается в «перепрыгивании» от клетки к клетке (горизонтальном распространении), тогда как дрожжевые прионы передаются вертикально — наследуются дочерними клетками при делении, переносясь в них с цитоплазмой. Стабильно передаваясь в линии дрожжевых клеток (как лабораторных, так и — в некоторых случаях — диких) и влияя на их фенотип, дрожжевые прионы представляют собой новую, белковую, форму гена — отступление от одного из правил центральной догмы молекулярной биологии (или дополнение к ней, кому как нравится). Если сравнивать дрожжевой прион с геном, то штаммовые варианты приона подобны аллелям этого гена. Только разница между такими «аллелями» не в последовательности мономеров носителя (в данном случае — белка), а в его пространственной укладке.
Все это позволяет изучать генетические свойства амилоидных полимеров в дрожжах — как они передаются и как их пространственная структура выполняет миссию носителя информации, сообщая новые черты клеточному фенотипу. В свое время ответы на подобные вопросы стали одними из самых фундаментальных открытий в исследованиях «классического» гена. Многие вопросы генетики дрожжевых прионов уже довольно хорошо изучены, но не исключено, что важные открытия еще впереди.
Лаборатория молекулярной генетики Института биохимии им. А. Н. Баха, в которой я работаю, издавна изучала процесс терминации трансляции у дрожжей, а в 1990-х годах, после открытия дрожжевых прионов, занялась и ими. Излюбленный объект лаборатории — прион [PSI+], наследуемый признак, в основе которого лежит амилоидная форма белка Sup35 (дрожжевого фактора терминации трансляции). Дрожжи, несущие в себе прион [PSI+], не особенно от него страдают, зато могут помочь нам в изучении тех прионных болезней, от которых страдаем мы. В клетках с генами, «поломанными» некоторыми типами мутаций, [PSI+] помогает восстанавливать их работу (рис. 4), в определенных условиях даже превращаясь в жизненно необходимый для дрожжей признак [6].
![Рис. 4. Механизм работы приона [PSI+] как полезного для дрожжей признака (Природа» №5, 2016)](https://elementy.ru/images/eltpub/infektsionnost_amiloidov_04_958.jpg)
Рис. 4. Механизм работы приона [PSI+] как полезного для дрожжей признака [6]. Белок Sup35 в норме прерывает трансляцию, останавливая рибосому на стоп-кодонах (слева). Коррумпированный полицейский, символизирующий прионную форму белка Sup35, перестает выполнять свою работу, позволяя машине-рибосоме проехать на запрещающий знак, преждевременный стоп-кодон, и синтезировать полноразмерный продукт гена Ade1 — фермент пути синтеза аденина. Благодаря этому клетка восстанавливает способность синтезировать аденин, потерянную в результате мутации
[PSI+] — один из наиболее изученных прионов, для него описано много штаммовых вариантов. Их принято условно делить на «сильные», хорошо связывающие мономерную форму Sup35 в амилоид, и «слабые», полимеризующие Sup35 менее эффективно, оставляя в клетке больше растворимой формы этого белка. Известно, что эффективность полимеризации Sup35 зависит от того, насколько хорошо идет фрагментация его фибрилл. Изучая амилоидные фибриллы, полученные из белка Sup35 in vitro, ученые обнаружили, что так называемое амилоидное ядро фибриллы — плотно уложенная, богатая β-слоями центральная ее часть — у «сильных» вариантов имеет меньший размер, чем у «слабых». В результате фибриллы, соответствующие «сильной» конформации, оказываются более ломкими и лучше фрагментируются, образуя большее количество свободных концов для полимеризации и тем самым повышая ее скорость.
Помимо приона [PSI+], белок Sup35 может образовывать также и ненаследуемый амилоид, который впервые описали 10 лет назад в нашей лаборатории [7]. Мы знаем, что этот амилоид массово образуется в клетках при сверхпродукции белка Sup35 и что он не может стабильно поддерживаться в клеточной линии, однако не представляем, в чем заключается его отличие от прионных амилоидов из того же самого белка. Почему одни амилоиды хорошо наследуются, а другие — нет, возникая в каждой дрожжевой клетке автономно? Многие данные наводили на мысль, что ненаследуемый амилоид должен иметь какую-то особую конформацию и, по всей видимости, особенно большой размер амилоидного ядра, что делает этот амилоид еще более слабым полимеризатором растворимого Sup35, чем самый «слабый» штамм приона [PSI+].
Ненаследуемый амилоид Sup35
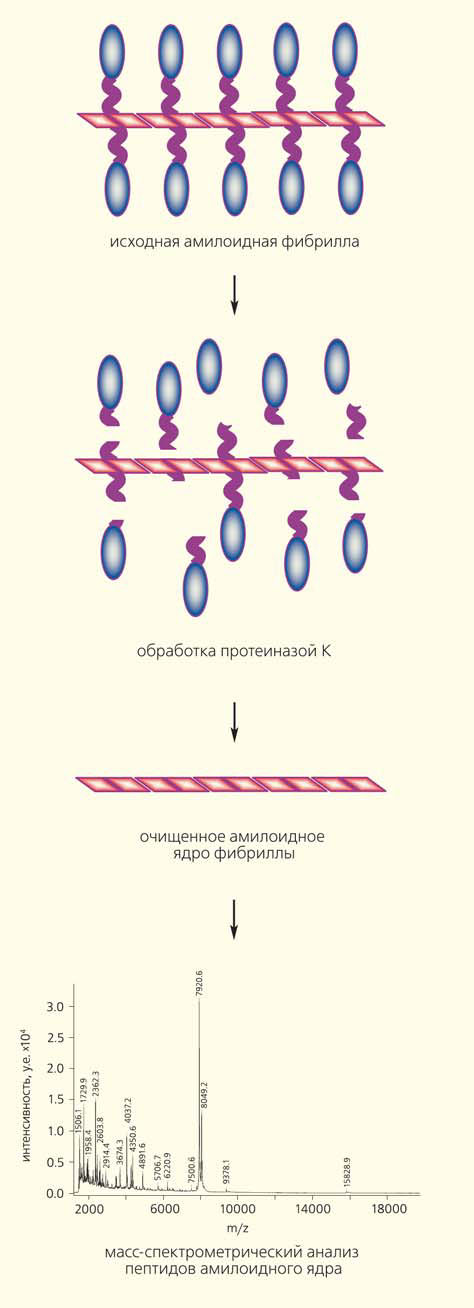
Рис. 5. Схема эксперимента по определению размера амилоидного ядра фибриллы
Амилоидное ядро устойчиво к действию протеаз, а потому мы можем определить его размер, расщепив протеиназой К все боковые, более «мягкие» части амилоидной фибриллы (рис. 5).
Некоторая проблема состояла в том, что амилоиды, выделяемые из дрожжей, прежде не получалось в достаточной мере очистить для такого анализа. Поэтому их приходилось «размножать», используя в качестве затравки для полимеризации in vitro высокоочищенного рекомбинантного Sup35, экспрессированного в Escherichia coli. Нам удалось усовершенствовать методику выделения амилоидных фибрилл из дрожжевых клеток, что сделало возможным непосредственный анализ их ядер, устойчивых к протеазам. Сравнив размер амилоидных ядер в препаратах с ненаследуемым амилоидом, а также с разными штаммовыми вариантами приона, мы вынуждены были отбросить первоначальную гипотезу (рис. 6).
Общая закономерность, касающаяся различий «сильного» и «слабого» вариантов приона, в эксперименте подтвердилась: амилоидное ядро «сильного» варианта меньше. Что же касается ненаследуемого амилоида, то его препарат содержал широкий спектр фрагментов Sup35, устойчивых к протеазам, размером от 3 до 8 кДа — от очень маленьких до весьма больших (но все равно не таких больших, как ожидалось). Все они имели одинаковое начало — N-конец белка Sup35. Таким образом, содержимое этого препарата соответствовало своеобразному «облаку» вариантов — смеси самых разных амилоидных конформаций, фибрилл с различной степенью ломкости, похожих на фибриллы «сильного» и «слабого» приона, а также промежуточных прионных вариантов.
Этот результат сам по себе не позволяет понять, существует ли это «облако» вариантов во всех клетках, из которых был выделен препарат амилоида, или же каждая клетка несет в себе какую-то одну, собственную индивидуальную укладку, а гетерогенная популяция клеток в сумме дает совокупность укладок.
![Рис. 6. Сравнение размеров амилоидного ядра ненаследуемого амилоида белка Sup35 и вариантов приона [PSI+] (Природа» №5, 2016)](https://elementy.ru/images/eltpub/infektsionnost_amiloidov_06_982.jpg)
Рис. 6. Сравнение размеров амилоидного ядра ненаследуемого амилоида белка Sup35 и вариантов приона [PSI+]. Линии над последовательностью амилоидогенного домена Sup35 и под ней обозначают размеры участков белка, уложенных в амилоидное ядро
Последнюю возможность мы отвергли с помощью ряда генетических тестов, которые продемонстрировали лабильность (способность подстраиваться под изменчивые условия окружающей среды) ненаследуемого амилоида в индивидуальных клетках. В частности, мы выращивали дрожжи в условиях дефицита аденина в среде, что подталкивало их уменьшать клеточное количество мономерной формы Sup35 (связь полимеризации Sup35 с метаболизмом аденина у дрожжей см. на рис. 4). Большинство клеток смогли переключиться в состояние, когда Sup35 полимеризуется в амилоид более эффективно. Это можно увидеть, например, по способу сборки фибрилл в макроскопические агрегаты: в нормальных условиях клетки несут в себе один амилоидный «комок» или «тяж», тогда как при дефиците аденина большинство клеток содержат множество центров агрегации фибрилл (рис. 7). В клетках с многочисленными мелкими агрегатами преобладают более ломкие конформации фибрилл, эффективно полимеризующие белок, а в клетках с единичными, более крупными агрегатами — более прочные и инертные фибриллы. Количественное соотношение разных конформаций в «облаке» сместилось в сторону тех из них, которые соответствуют «сильным» вариантам приона. Сама возможность такого смещения в значительной части клеток свидетельствует о присутствии в каждой из них полноценного «облака» конформаций, внутри которого возможна селекция. Кстати, это смещение оказалось еще и обратимым: добавив в среду нормальное количество аденина, селекцию амилоидных конформаций можно повернуть в противоположную сторону.

Рис. 7. Форма макроскопической агрегации амилоида позволяет отслеживать изменение его свойств под влиянием окружающей среды. В дрожжах, выращенных в среде с нормальным (слева) и минимальным количеством аденина, формы агрегации заметно различаются. Циановым цветом показаны амилоидные агрегаты из белка Sup35-GFP, желто-зеленым — автофлуоресценция клеток
Таким образом, мы выяснили, что изучаемое нами вещество очень гетерогенно по конформациям составляющих его фибрилл и может изменять свои свойства в зависимости от условий. Помимо картирования амилоидного ядра с помощью протеиназы, безусловно, существуют и другие способы изучать укладку белка в амилоид, однако есть основания полагать, что с ненаследуемым амилоидом они также не покажут четкого отличия от прионов. Если причина неинфекционности нашего амилоида заключается не в однородно большом амилоидном ядре, как мы предполагали, то, возможно, конформация амилоида и вовсе не играет в этом вопросе решающей роли. Основной вклад в неинфекционность амилоида может вносить какой-то иной фактор, и кто знает, не воспользуемся ли мы им в будущем для борьбы с прионными болезнями, делая их менее заразными? Если похожее «облако» вариантов обнаружится и у человеческих амилоидов, не сможем ли мы действовать на него таким образом, чтобы свести на нет то вредоносное влияние, которое амилоидные агрегаты оказывают на клетки мозга? Сможем ли мы найти такие лекарства, которые блокировали бы полимеризацию всех штаммовых вариантов в «облаке», не позволяя отдельным из них проявить резистентность и резко снизить эффективность всего лечения? Открытие, сделанное на одноклеточных и совсем не страдающих от нейродегенерации организмах, может положить начало большим свершениям.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (проект 14-04-00073), а также Программы по молекулярной и клеточной биологии Российской академии наук.
Литература
1. Tycko R., Wickner R. B. Molecular structures of amyloid and prion fibrils: consensus versus controversy // Acc. Chem. Res. 2013. V. 46. P. 1487–1496. DOI: 10.1021/ar300282r.
2. Asante E. A., Smidak M., Grimshaw A. et al. A naturally occurring variant of the human prion protein completely prevents prion disease // Nature. 2015. V. 522. P. 478–481. DOI: 10.1038/nature14510.
3. Chimon S., Shaibat M. A., Jones C. R., Calero D. C., Aizezi B., Ishii Y. Evidence of fibril-like β-sheet structures in a neurotoxic amyloid intermediate of Alzheimer’s β-amyloid // Nat. Struct. Mol. Biol. 2007. V. 14. P. 1157–1164. DOI: 10.1038/nsmb1345.
4. Goedert M. Neurodegeneration. Alzheimer’s and Parkinson’s diseases: The prion concept in relation to assembled Aβ, tau, and α-synuclein // Science. 2015. V. 349. P. 1255555. DOI: 10.1126/science.1255555.
5. Khurana V., Lindquist S. Modelling neurodegeneration in Saccharomyces cerevisiae: why cook with baker’s yeast? // Nat. Rev. Neurosci. 2010. V. 11. P. 436–449. DOI: 10.1038/nrn2809.
6. Liebman S. W., Chernoff Y. O. Prions in yeast // Genetics. 2012. V. 191. P. 1041–1072. DOI: 10.1534/genetics.111.137760.
7. Salnikova A. B., Kryndushkin D. S., Smirnov V. N., Kushnirov V. V., Ter-Avanesyan M. D. Nonsense suppression in yeast cells overproducing Sup35 (eRF3) is caused by its non-heritable amyloids // J. Biol. Chem. 2005. V. 280. P. 8808–8812. DOI: 10.1074/jbc.M410150200.
* Со статьей «Прионные и неприонные амилоиды: определяет ли конформация разницу в инфекционности?», участвовавшей в научно-популярном конкурсе «Био/мол/текст»-2015 в номинации «Своя работа», можно ознакомиться на сайте «Биомолекула», посвященном молекулярным основам современной биологии и практическим применениям научных достижений в медицине и биотехнологии. По договоренности с организаторами конкурса мы публикуем переработанный вариант этой статьи. — Примеч. ред.
** Подробнее см.: Кушниров В. В. Прионы и амилоиды: ключевые свойства и роль в природе // Природа. 2014. № 3. С. 37–43.











Рис. 1. Фибриллы Aβ-амилоида под электронным микроскопом (слева) и схематическое изображение параллельной и антипараллельной кросс-β-структуры амилоидной фибриллы [1]