Юрские динозавры Забайкалья
Владимир Алифанов, Сергей Савельев
«Природа» №5, 2016
 Владимир Рудольфович Алифанов — кандидат биологических наук, старший научный сотрудник Палеонтологического института им. А. А. Борисяка РАН, специалист в области морфологии, филогении и систематики ископаемых пресмыкающихся. |
 Сергей Вячеславович Савельев — доктор биологических наук, заведующий лабораторией развития нервной системы Института морфологии человека РАМН. Область научных интересов — эмбриология и патоморфология нервной системы животных и человека, механика развития, зоопсихология и палеоневрология. |
Открытию динозавров в Кулинде (верхняя юра) в Забайкалье была посвящена первая часть статьи (см. «Природу» № 4, 2016, с. 35–44). В ней также рассказывалось о политике местных ученых, которая привела к формированию двух параллельных программ изучения, о противоречиях и просчетах в изучении ископаемых остатков, о конкретных видах из нового местонахождения и об особенностях эволюции групп, к которым они принадлежат. Однако тема этим не исчерпывается. Важно обсудить проблему возраста отложений, откуда извлечены ископаемые остатки древних пресмыкающихся, оценить строение необычных покровных образований, которые встречаются в местонахождении, и гипотезы о происхождении пера. Находки из Кулинды дают также повод задуматься над проблемами классификации крупнейших таксономических групп динозавров.
Неизвестная страница летописи
Состав комплекса ископаемых из конкретного местонахождения позволяет охарактеризовать его в качестве или типичного, или уникального. В первом случае данные можно использовать как аргумент для определения стратиграфического возраста вмещающих органические остатки пород, а во втором появляется возможность говорить о неизвестной вехе ископаемой летописи. Такая веха может стать основой для региональных или более широких фаунистических, биостратиграфических и палеобиогеографических сравнений в дальнейшем. Однако чаще всего тот или иной комплекс интегрировать в существующие стратиграфические схемы даже узкорегионального масштаба непросто. Неопределенность — следствие ограниченного количества данных. Именно неопределенный характер носили до недавнего времени находки из отложений укурейской свиты.
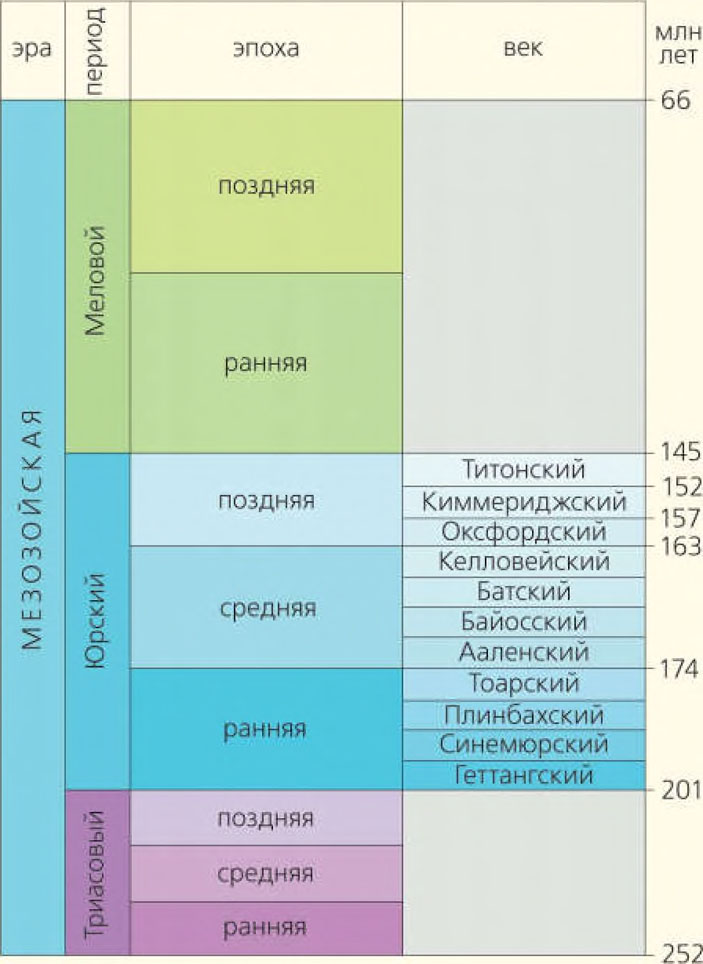
Ранее укурейскую свиту датировали и ранним мелом, и поздней юрой в целом или только ее началом. Есть результаты радиоизотопного датирования K-Ar-методом. Они показывают возможность образования укурейских отложений в интервале от 169 до 144 млн лет назад, т. е. в стратиграфическом диапазоне от средней юры до начала раннего мела, что практически ничего не добавляет к имеющейся палитре мнений.
Решить весьма старый научный спор помогают находки динозавров из Кулинды, которые принадлежат, как говорилось в первой части, представителям семейств гипсилофодонтид и джехолозаврид (орнитоподы), а также нквебазаврид (орнитомимозавры). Для более полной характеристики герпетофауны, установленной на сегодня в данном местонахождении, следует отметить еще один ее элемент. Это титанозавры (Titanosauria), относящиеся к группе завропод (Sauropoda) в составе ящеротазовых динозавров (Saurischia). Первые титанозаврообразные ящеры появились в самом конце юрского периода и достигли большого разнообразия в мелу. В Кулинде находки этой группы представлены покровными окостенениями — остеодермами — в виде характерных округлых пластин, пронизанных отверстиями. В ранних сборах этих материалов не было, но оказалось возможным сделать определение по еще не опубликованным фотографиям образцов из сборов 2014 г.
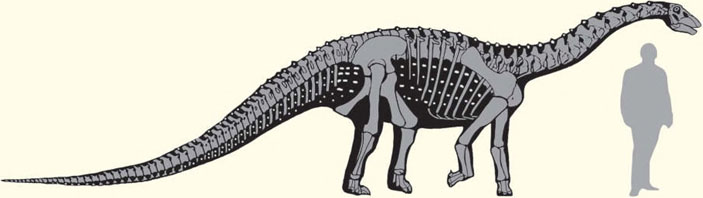
Контурная реконструкция внешнего вида (с вписанным в нее скелетом) архаичного титанозаврообразного завропода. Белыми пятнами показано предположительное положение остеодерм. Здесь и далее рисунки В. Р. Алифанова
Теперь можно кратко обсудить датировку слоев укурейской свиты (содержащих местонахождение Кулинда), используя определения состава фауны динозавров. Тому, что они образовались в течение раннего мела, противоречат находки гипсилофодонтид, которые отсутствовали в то время на Азиатском континенте. Аргумент против отнесения к средней юре и даже к началу поздней юры — остатки таких эволюционно молодых групп, как титанозавры, орнитомимозавры и джехолозавриды. Итог представляется следующим: комплекс кулиндийских динозавров, с одной стороны, уникален, а с другой — носит переходные черты от типично юрских к типично раннемеловым. Скорее всего, он существовал в самом конце поздней юры, т. е. в течение титонского века.
Если не брать в расчет отдельные и редкие находки, то комплексов наземных позвоночных такого возраста в Центральной Азии нет. Таким образом, главное значение материалов из местонахождения Кулинда — это частичная ликвидация пробела в палеонтологической летописи большого региона, приходящегося на рубеж юры и мела. Интересно, что коллеги из параллельного проекта, связанного с изучением кулиндинской фауны, не смогли склониться ни к одному из конкретных вариантов оценки возраста укурейской свиты и предпочли остаться в рамках неопределенной радиоизотопной датировки, отказавшись без объяснения причин только от раннемеловой части показанного ею интервала.
Меньше всего противоречий в рамках двух проектов содержится в реконструкции условий жизни кулиндийских динозавров.
Остатки ящеров погребены в озерных отложениях с характерной для них однородностью и мелкозернистостью. На характеристики водоема и состав осадка влияла периодическая вулканическая активность в районе образования местонахождения. Сохранение покровных структур вместе с костями указывает на восстановительные условия среды в осадке и быстрое захоронение. Редкость сочлененных элементов скелета свидетельствует о том, что тела погибших животных перед погребением подвергались мацерации (т. е. разрушению мягких тканей) и недалекому переносу (вероятно, из береговой зоны) к месту захоронения. Озеро было обширным, местами мелководным. Его берега частично зарастали мхами и хвощами, а кое-где — крупноствольными деревьями.
Растительность, несомненно, давала убежище и пищу двуногим орнитоподам. Кормом служили как вегетативные (стебли, листья), так и генеративные (т. е. связанные с размножением — соплодия, семена) части голосеменных (кейтониевых, чекановских, гинкговых и др.) растений, которые доминировали в юрское время. Вряд ли динозавры обходили вниманием беспозвоночных животных, особенно насекомых и их личинок, находки которых либо обнаружены в самой Кулинде, либо известны из отложений укурейской свиты и ее возрастных аналогов в других районах Забайкалья. Для хищных динозавров объектом охоты кроме насекомых, несомненно, были рыбы, земноводные, ящерицы и другие мелкие позвоночные, о которых нам лучше известно по находкам из упоминавшегося в первой части статьи местонахождения Аулие (Каратау) в Южном Казахстане.
Проблема происхождения пера
Ключевая идея ныне действующей системы взглядов на эволюцию динозавров — родство теропод (хищных динозавров) и птиц. Она многократно обсуждалась и ранее, но особенную убедительность приобрела в связи с открытиями, начавшимися примерно четверть века назад в Северо-Восточном Китае. Там, особенно в провинции Ляонин, довольно широко распространены континентальные отложения раннемелового возраста, в которых время от времени обнаруживаются скелеты представителей разных групп теропод с остатками покровов в виде удлиненных придатков, в том числе перьев птичьего типа. Подобные открытия привели к появлению концепции оперенных динозавров, а также актуализировали вопросы о первоначальной функции и происхождении пера.
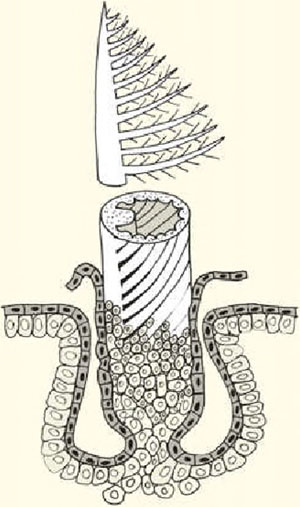
Схематическое изображение пера современных птиц. Основание, или очин, всех модификаций пера — это полая цилиндрическая структура
Перо — это особый тип покровных образований. Оно имеет цилиндрическое основание, или очин, переходящее в стержень, поддерживающий опахало. Последнее составлено из сцепленных друг с другом бородок. Иногда признаки типичного строения пера меняются. Например, вместо опахала может формироваться пуховой, щетинковидный или нитевидный вариант. Большое морфологическое разнообразие перьев определяет и их связь с разными функциями: защитной, демонстрационной, механорецепторной, термоизоляционной и локомоторной. Если первые три функции имеют дополнительное значение, то последние две — основные, поскольку связаны с машущим полетом. Новые исследования показывают, что некоторые динозавры также летали с помощью оперенных крыльев, но, в отличие от птиц, их полет носил планирующий характер. Что касается других функций динозавровых перьев в сравнении с птичьими, то сомнение вызывает только способность к эффективной термоизоляции, поскольку в специализированном виде она, скорее всего, возникла с развитием пухового слоя под кроющим оперением. Такое сочетание покровных элементов у динозавров выявить пока не удалось.
Следует также сказать о проблеме гомологии пера. Традиционно его происхождение ведут от чешуи рептилий, поскольку на самом начальном этапе зачаток пера оформляется как чешуя пресмыкающихся. Далее развитие приводит к погружению вглубь кожи краев чешуйки, где они захватывают внутрь себя участок мезодермы с сосудами, образуя так называемый сосочек. Через него затем будут поддерживаться ростовые процессы. Верхняя часть внутреннего слоя будущего пера дифференцируется на серию выступающих внутрь гребней, участвующих в образовании бородок. Очевидно, что на пути эволюционного становления пера произошел ряд эмбриональных и морфологических преобразований. Однако их сущность и последовательность возникновения остаются загадкой и обычно объясняются в соответствии со стадиями, наблюдаемыми при формировании пера у современных птиц.
Палеонтологические данные пока что не вносят ясности в этот вопрос, хотя новые материалы достаточно разнообразны и в некоторых случаях весьма примечательны. Например, у ляонинских теропод отмечается около 10 морфотипов перообразных структур. Большая их часть имеет стержень и напоминает пух или перо птичьего типа, а меньшая включает структуры, которые иногда называют нестержневым пером. В числе последних преобладают нитевидные или волосовидные образования, получившие название «протоперо». Еще один нестержневой вариант — это пучки волокон с общим основанием; его иногда разделяют на два типа: с параллельными или с расходящимися нитями. Связь стержневых и нестержневых структур друг с другом остается невыясненной. Длинные нестержневые покровные производные встречены не только у теропод, но также и у двух групп птицетазовых динозавров: пситтакозаврид [1] и гетеродонтозаврид [2]. У первых они толще и длиннее, чем у теропод, а у вторых выглядят как типичные протоперья.

Кости передней конечности с прилегающим к ним полем покровных образований и прорисовка моно- и полищетинковых вариантов строения отдельных щетинковых чешуй. Двуцветная окраска некоторых чешуй свидетельствует о возможной погруженности части чешуи в дерму. Здесь и далее фото С. В. Савельева
Понятно, что обозначение всего многообразия удлиненных покровных образований «перьями», обычное в нынешней литературе и принимаемое также авторами описания «кулиндадрома», ведет к размыванию строгого определения пера как структуры, имеющей прежде всего стволовидное цилиндрическое основание. В указанном, предельно широком, использовании этого термина, несомненно, отражается отмеченное выше отсутствие ясной концепции становления настоящего пера, а также неоднозначность представлений о том, какую роль в этом процессе могли играть (если играли) те или иные варианты древних пероподобных структур.
Между тем пролить свет на некоторые тайны происхождения пера и, в первую очередь, его стволовидного основания, помогает изучение строения кожных производных орнитопод из Кулинды. Они представлены небольшими уплощенными чешуями (шириной от 2 до 6,5 мм), внешне напоминающими по строению типичную чешую пресмыкающихся. Однако от их внешнего края отходит серия отдельных полосок, или щетинок, шириной 0,1–0,3 мм и длиной до 70 мм. Такого рода образования мы предложили называть щетинковой чешуей [3–5]. Среди них встречаются варианты с 3–8 щетинками. Есть и монощетинковая чешуя, соответствующая протоперу. В целом щетинковые чешуи напоминают нестержневые покровные придатки теропод, но отличаются тем, что соседние нити (щетинки) уплощены и немногочисленны. Данные об изолированном захоронении щетинковых чешуй, о разной длине щетинок, иногда об их цветовом контрасте с основаниями чешуй наводят на предположение, что покровные образования у кулиндийских орнитопод обладали пролонгированной фазой роста, были частично погружены в дерму и могли выпадать при линьке. Такие признаки свойственны и птичьему перу. Но у щетинковой чешуи из Кулинды нет стержневого основания, составляющего, как уже было сказано, ключевую характеристику настоящего пера.

Поле удлиненных и спутанных покровных придатков передней конечности, принадлежащих предположительно кулиндаптериксу, и схема преобразования простой чешуи в щетинковую, а щетинковой чешуи — в пухообразную структуру. Пространственное изменение строения щетинковой чешуи путем ее трансформации в трубковидную структуру может быть принято в качестве модели для происхождения настоящего пера и установления гомологии его основных элементов
Моно- и полищетинковые модификации нестержневых перьев, которые отмечаются также и у кулиндийских орнитопод, наводят на мысль, что это морфологические вариации особого типа — его нельзя свести к простой чешуе пресмыкающихся, но нельзя и принять за настоящее перо. Иначе говоря, нестержневые образования представляют собой некую промежуточную стадию между чешуей пресмыкающихся и пером птиц. Такой взгляд на обсуждаемые структуры в литературе отчетливо еще не прозвучал.
Остается вопрос, какой именно нестержневой вариант стал основой для происхождения настоящего пера. До сих пор предпочтение отдавалось монощетинковому протоперу, которое стало известно раньше щетинковой (т. е. полищетинковой) чешуи. Нам же кажется, что перу птичьего типа предшествовал не первый, а второй вариант.
В выборе гипотезы существенно то, что среди вариаций щетинковых чешуй отмечается разная степень изгибания ее базальной части вдоль длинной оси, вплоть до образования полукольца в поперечном сечении. Логично предположить, что дальнейшее усиление этого процесса в ходе эволюции могло привести к формированию трубковидной структуры, соответствующей очину типичного пера. В этом случае щетинки, образуемые чешуями такого типа, могут быть с большей вероятностью гомологизированы с бородками пухового пера. Гипотеза происхождения птичьего пера из протопера требует гораздо большего числа допущений, за которыми реальных морфологических данных пока нет.
Поиск «истинных» динозавров
Находки динозавров из Кулинды интересны не только тем, что могут пролить свет на решение ряда конкретных проблем в изучении некоторых групп птицетазовых динозавров и теропод, но и тем, что позволяют уточнить общие представления о путях эволюции динозавров. В качестве ключевого здесь можно выделить вопрос о том, каким образом современные данные об удлиненных покровных структурах у динозавров могут влиять на традиционные представления об основных направлениях филогенетической дифференциации этой группы, а следовательно, и на интерпретацию родственных отношений между ними. И в этом случае между двумя параллельными проектами пролегла разделительная линия.
Авторы описания «кулиндадрома» заключили, что нестержневые перья в виде протопера имели широкое распространение у динозавров, включая их самых архаичных представителей. Такой вывод означает не что иное, как распространение концепции оперенных динозавров, которая в первоначальном виде имела отношение только к тероподам, на всех динозавров. В наших работах также сделано похожее и напрашивающееся уже по итогам более ранних находок в Китае заключение [3–5], которое, однако, дальше птицетазовых динозавров не распространялось. За бортом этой концепции, с нашей точки зрения, следовало бы оставить прозавропод и завропод (их обычно объединяют в группу завроподоморф), для которых нет и намека на наличие каких-либо перообразных покровных придатков. Различия в оценках обнажают не сразу заметные противоречия в действующей системе представлений об эволюции макротаксонов динозавров. А на них стоит обратить внимание.

Филогенетические связи архозавров по А. Ромеру [6]. На схеме предками архозавров и птиц указаны текодонты. Динозавры представлены двумя независимыми ветвями без упоминания прозавропод. Такая схема предполагает, что признаки сходства высших архозавров (т. е. птерозавров и динозавров) и птиц возникли независимо друг от друга
Здесь следует вспомнить, что до середины 1980-х годов динозавров нередко воспринимали как сборную группу. Сомнительной считалась непосредственная связь птицетазовых и ящеротазовых динозавров, прозавропод и завропод, что отчасти отражает схема А. Ромера [6], хорошо известного в России морфолога и палеонтолога. Впрочем, строгих доказательств филогенетического родства названных групп нет и сейчас. С нашей точки зрения, большое сомнение вызывает даже естественность объединения завропод и теропод в рамках таксона Saurischia (ящеротазовые динозавры), поскольку так называемая ящеротазовость (имеется ввиду ящерицетазовость), которая легла в основу характеристики самой большой подгруппы динозавров, есть по сути лишь общая примитивная (плезиоморфная) особенность, не указывающая на специфическое родство ее носителей. Такое объединение не соответствует современным принципам систематики и филогении, подразумевающим выделение и сближение групп на основе характерных для них черт специализации.
Вместе с тем неучтенной характеристикой, а точнее, адаптацией системного уровня, т. е. затрагивающей широкий спектр морфологических преобразований (и заслуживающей внимания как возможный индикатор родства), можно считать формирование среди части динозавров способности к двуногому передвижению. Из-за отсутствия этой адаптации и сопутствующего ей комплекса признаков четвероногие завроподы, как нам представляется, должны занимать на филогенетическом древе динозавров более базальное положение, а вот тероподы и птицетазовые динозавры, в составе которых обычны облигатно-бипедальные формы, заслуживают исследования на предмет их близкого родства.

Тазовые кости (вид сбоку) некоторых групп «ящеротазовых» пресмыкающихся. В середине — брахиоилический таз прозавропода текодонтозавра (Thecodontosaurus), который сходен с тазом текодонта эупаркерии (Euparkeria). Закрытый таз завропода ферганозавра (Ferganosaurus) отличается развитием сложного крестца и признаком долихоилия, т. е. удлиненной подвздошной костью. Обозначения костей: Л — лобковая, П — подвздошная, С — седалищная
О таком родстве отчасти свидетельствуют данные строения таза. Например, для теропод и птицетазовых динозавров отмечается так называемая опистопубия, т. е. ориентация лобковых костей назад, как у птиц (что даже отмечено в названии второй группы динозавров). Не исключено, что для теропод исходной была не препубия (ориентация лобковых костей вперед), как у завроподоморф, а по крайней мере пропубия, которая означает промежуточное между пре- и опистопубией состояние. Важно и то, что у бипедальных динозавров таз часто не сплошной, а облегченный: у теропод — благодаря возникновению вентрального (нижнего) окна, называемого тироидным, а у птицетазовых динозавров — за счет утраты связи лобковых костей друг с другом. Все эти признаки прозавроподам и завроподам не свойственны, а их таз (судя по строению этой части скелета у представителей других групп динозавров и птиц) архаичен. Особенно примитивен он у прозавропод из-за отсутствия у них сложного крестца, связанного с наличием четырех и более тазовых позвонков и развитием удлиненной подвздошной кости (долихоилия). В соответствии с этим таз прозавропод иногда типизируют как брахиолический, т. е. имеющий укороченное строение подвздошных костей. Этот признак также соответствует тазу ящериц, текодонтов и многих более архаичных, чем динозавры, групп пресмыкающихся.
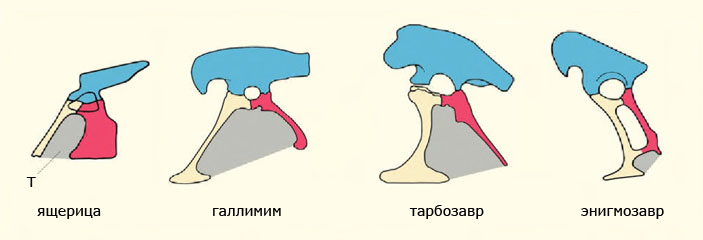
Фенестрированные с помощью тироидного окна тазы пресмыкающихся: брахиоилический таз ящериц и долихоилические варианты тазов теропод, представленные препубическим вариантом у галлимима (Gallimimus), пропубическим у тарбозавра (Tarbosaurus) и опистопубическим у энигмозавра (Enigmosaurus). Разнообразие строения таза теропод не допускает однозначной возможности определения их родственных связей в составе динозавров на основании так называемой ящеротазовости. Серым цветом обозначено тироидное окно (Т) между тазовыми костями
Связь теропод и птицетазовых динозавров устанавливается и по строению стоп, которые, в отличие от стоп завроподоморф, приобретают внешне симметричную форму из-за редукции первого и пятого пальцев вместе с увеличенными размерами третьего (среднего) пальца. Для двух групп также характерно пальцехождение в отличие от архаичного стопохождения у завропод, у которых к тому же произошла редукция числа элементов большинства пальцев, но не самих пальцев.
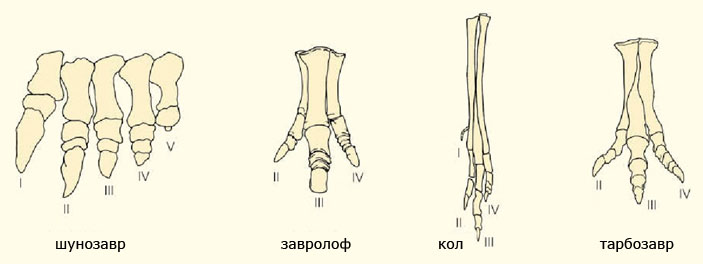
Стопы разных групп динозавров: завропода шунозавра (Shunosaurus), птицетазового динозавра завролофа (Saurolophus), альваресзавра кола (Kol) и теропода тарбозавра (Tarbosaurus). Короткие и широко расставленные пальцы завропода указывают на стопоходящий тип походки, а компактная и высокая стопа у остальных — на пальцехождение. Латинскими цифрами обозначены порядковые номера пальцев
Список признаков сходства птицетазовых динозавров и теропод невелик. Тем весомее представляется проявление тенденции к формированию у тех и у других перообразных структур, которые, заметим, присутствуют только у двуногоходящих (бипедальных) динозавров. Связь удлиненных покровных образований и способа походки выглядит и закономерно, и загадочно, а главное — плохо объясняется существующими теоретическими представлениями. Можно, например, пофантазировать и предположить, что особенности двуногой походки требовали более высокой физиологической активности и, соответственно, специфического типа линьки, которая охватывала бы не весь покров сразу, а только его отдельные участки, за счет чего снижался бы уровень биологической уязвимости (у линяющих целиком пресмыкающихся он обычно высок).
Подобные рассуждения позволяют подойти к решению проблемы общей характеристики динозавров. В рамках текущей системы взглядов, сложившейся во многом традиционно, существующее формальное определение выглядит недостаточным, поскольку касается таких признаков, как наличие сложного крестца в сочетании с несколькими специфическими чертами строения костей конечностей. Отсутствие ясной и исчерпывающей характеристики давно ставит перед исследователями проблему «истинных» динозавров (т. е. не сборной группы, а естественного таксона), в число которых, не исключено, пока включают группы и формы, лишь внешне динозаврообразные. Общая картина останется неясной и противоречивой, если не предположить, что в эволюции пресмыкающихся происходило независимое формирование набора сходных признаков у динозавров и у их близких и дальних родственников. С нашей точки зрения, проблема «истинных» динозавров пока не решена.
Интересно, что весьма большое количество черт сходства с динозаврами демонстрируют летающие ящеры (птерозавры). Исторически сложилось, что их относят к особой группе среди архозавров вне рамок собственно динозавров. Хотя динозаврообразность птерозавров не ускользнула от исследователей и не раз обсуждалась в литературе, сходство их с динозаврами признается вторичным. Однако если птерозавры — не динозавры, то почему динозаврами считаются, например, те же прозавроподы, лишенные сложного крестца?
Ситуацию могли бы прояснить новые ископаемые находки промежуточных звеньев эволюционного процесса. На первый взгляд, такое место между тероподами и завроподами занимают прозавроподы. Однако в отношении именно последней группы как раз и возникает больше всего сомнений в их принадлежности к динозаврам. Если придерживаться традиционной точки зрения, то следует признать, что один из главных признаков динозавров — сложный крестец — возникал в эволюции архозавров многократно и параллельно, что возможно. Но отнести эту независимость на счет динозавров — значит поставить под сомнение естественность этой группы. Странно, что при таком грузе нерешенных проблем не уделяется внимания существованию промежуточной между тероподами и птицетазовыми динозаврами группы. Это, по нашему мнению, так называемые альваресзавры (Alvarezsauria, в кладистических классификациях Alvarezsauroidea), которые были открыты в конце 1990-х годов в Аргентине и Монголии.
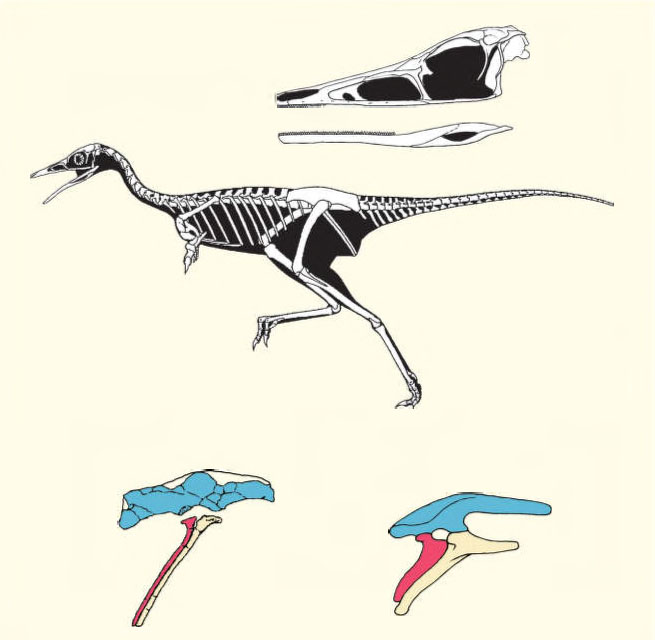
Реконструкция скелета и черепа альваресзавра шувууйи (Shuvuuia) из верхнего мела Монголии, а также сравнение его таза с тазом птицетазового динозавра — стегозавра (справа), конструкция которого имеет сходное строение. Скелет альваресзавра сочетает признаки теропод и птицетазовых динозавров
Первые находки альваресзавров носили фрагментарный характер. Поэтому аргентинские материалы позволили исследователям допустить родство альваресзавров с орнитомимозаврами, а монгольские — с архаичными птицами, но более продвинутыми, чем археоптерикс. Потом, когда появились новые данные из Аргентины и Монголии, удалось установить, что находки на двух удаленных друг от друга континентах принадлежат общей группе. Ее все же отнесли к тероподам. Но ящеры эти очень странные. Таз у них имеет такое же строение, как у птицетазовых динозавров, а вот стопа по компактности действительно уклоняется в сторону теропод, хотя и отличается некоторыми уникальными деталями строения. Череп внешне похож на тероподовый, а нижняя челюсть имеет признаки строения, характерные для птицетазовых динозавров. Для альваресзавров не установлены брюшные ребра, которых, может быть, и не было, как у птицетазовых динозавров. И еще: у альваресзавров, возможно, кисть была пятипалой, если приплюсовать к трем известным пальцам два шиповидных образования в области кисти, установленных при описании цератоника (Ceratonykus) из верхнего мела Монголии. Пять пальцев передней конечности обычны для птицетазовых динозавров, но не для теропод, у которых в редких случаях сохраняются рудименты четвертого пальца. С тероподами альваресзавры могут быть сближены лишь с большой натяжкой, но их можно рассматривать как близких родственников птицетазовых динозавров [7], коль скоро у них тазы сходного типа.
Пример с альваресзаврами хорошо дополняется данными о покровах. Вместе они показывают, что выделение крупных таксономических групп динозавров и реконструкция их родственных взаимоотношений имеет немало теоретических условностей, а значит, и перспективы для существенной коррекции.

Общепринятая филогенетическая схема основных таксонов динозавров. Она демонстрирует обособленное положение птицетазовых динозавров, связь с динозаврами прозавропод и родство птиц с хищными динозаврами (в соответствии с тероподовой гипотезой)
Учесть наши сомнения в отношении текущей схемы родственных взаимоотношений динозавров можно в виде филограммы, отражающей родственные связи основных групп архозавров. Такая графическая картина позволяет наглядно продемонстрировать представление об «истинных» динозаврах и о том, кем были их близкие или дальние родственники. Теперь сомнения в принадлежности к динозаврам можно отнести не только к прозавроподам, но и к завроподам. Если так, то завроподы, как и птерозавры, вполне могут принадлежать к группе самых динозаврообразных архозавров. На этом фоне прозавроподы, скорее всего, окажутся близкими родственниками не завропод (или в целом динозавров), а текодонтов (на филограмме они не показаны), среди которых также много динозаврообразных групп и форм.
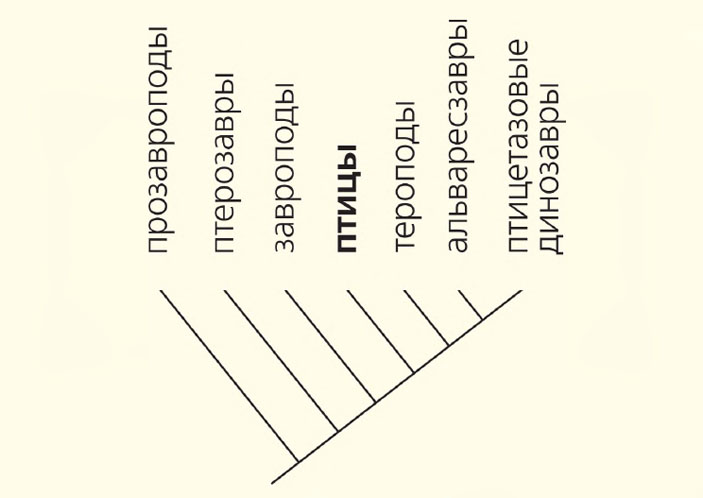
Филогенетическая схема, основанная на новой гипотезе родственных связей высших архозавров, с указанием положения птиц. Введение в схему продвинутых морфологически птерозавров оттеняет сомнительность включения в состав динозавров прозавропод. Предположение о происхождении птиц и динозавров от общего неспециализированного предка позволяет обособить истинных динозавров от их динозавроподобных родственников, т. е. от завропод и прозавропод. Эти родственники динозавров демонстрируют тенденцию к приобретению некоторых (проявляющихся мозаично) динозавровых признаков, тогда как у собственно динозавров происходил процесс приобретения сходства с птицами
В итоге можно зафиксировать важное и давно высказанное предположение о существовании в эволюции архозавров процесса динозавризации, т. е. приобретения параллельного с «истинными» динозаврами определенного набора признаков. Интересно, что основное направление динозавризации связано с уходом от образа архаичной рептилии, которая во время движения изгибает свое тело вбок, что помогает ей переставлять короткие и расставленные в сторону конечности. В эволюции архозавров боковые изгибы тела сократились, а задние конечности испытывали тенденцию к удлинению вместе с ориентацией коленного сустава вперед, что означает заведение ног под туловище, как у млекопитающих. Это сделало походку более экономной с энергетической точки зрения и, вероятно, более быстрой, что в условиях открытых и выровненных ландшафтов имело существенное значение. На этом эволюционном пути наметилось отставание развития передних конечностей. С появлением сложного крестца, как оказалось, можно избавиться от участия передних конечностей в передвижении, что, похоже, смогли сделать настоящие динозавры.
Несовершенство традиционной базовой схемы родственных взаимоотношений динозавров не может не проявиться в таком интригующем для современных исследователей вопросе, как родство динозавров и птиц. Он сложен и многогранен и вряд ли может быть до конца прояснен даже с помощью специального и самого подробного разбора. Сегодня уже можно смело расстаться с представлениями о происхождении птиц от каких-то архаичных и четвероногих (по способу передвижения) пресмыкающихся. Однако и преобладающая ныне гипотеза, выводящая птиц от теропод, не кажется столь бесспорной, насколько это может внушаться фактом ее практически всеобщего признания.
Процесс становления тех или иных черт птичьего плана строения у динозавров не раз отмечался исследователями. Нашел он отражение в трудах Р. Барсболда [8] и С. М. Курзанова [9], которые данный процесс стали называть орнитизацией. Проанализирован он этими учеными на примере проявлений орнитизации только у теропод. Но теперь, благодаря новым содержательным данным о перообразных покровах, в сферу анализа могут быть включены и бипедальные птицетазовые динозавры. И здесь возникает возможность сделать предположение о происхождении птиц, которое еще никем не высказывалось.
Совсем не исключено, что все способные к бипедальной походке динозавры прошли с птицами параллельный путь эволюции от общего неспециализированного предка. При этом представители некоторых линий теропод (особенно троодонтий, дромеозавров и овирапторозавров) продвинулись на пути орнитизации дальше других, достигнув наибольшего внешнего сходства с самыми архаичными птицами. Если так, то именно птицы позволяют нам понять, кто был «истинным» представителем динозавров, а кто — родственной им группой.
***
Материалы по оперенным птицетазовым динозаврам из Забайкалья — всего лишь звено в цепи необычных находок, ставших известными в последнее годы. Все они ведут к тем или иным новым теоретическим заключениям. С их помощью можно попытаться защитить традиционную систему взглядов, а можно попробовать вскрыть ее слабости. И в том, и в другом случае необходима упорная исследовательская работа, с учетом новых данных, подходов и промежуточных предположений. Чтобы глубже проникнуть в тайну происхождения и эволюции динозавров, нужны также и новые открытия, подобные тем, что принесло местонахождение Кулинда.
Работа поддержана Российским фондом фундаментальных исследований (проекты 13-05-00302 и 16-05-00408) и Акционерным некоммерческим обществом «Институт биомедицинских проблем» (грант «Палеонтология позвоночных»).
Литература
1. Mayr G., Peters D. S., Plodowski G. Bristle-like integumentary structure at the tail o f the horned dinosaur Psittacosaurus // Naturwissenschaften. 2002. V. 89. № 8. S. 361–365.
2. Zheng X., You H., Xu X. et al. An Early Cretaceous heterodontosaurid dinosaur with filamentous integumentary structures // Nature. 2009. V. 458. № 7236. P. 333–336.
3. Савельев С. В. Алифанов B. P. Новый тип кожных дериватов у птицетазовых динозавров из поздней юры Забайкалья // Докл. РАН. 2014. Т. 456. № 2. С. 251–253.
4. Алифанов В. Р., Савельев С. В. Два новых птицетазовых динозавра (Hypsilophodontia, Ornithopoda) из поздней юры России // Палеонтол. журн. 2014. № 4. С. 72–82.
5. Алифанов В. Р., Савельев С. В., Терещенко Е. Ю. и др. Строение кожных покровов у птицетазовых динозавров (Hypsilophodontia, Ornithopoda) из поздней юры Забайкалья // Палеонтол. журн. 2014. № 5. С. 72–80.
6. Romer A. Sh. The Vertebrate Story. Chicago, 1959.
7. Алифанов B. P. Инфраотряд Alvarezsauria // Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. 4.2 / Отв. ред. Е. Н. Курочкин, А. В. Лопатин. М., 2012. С. 325–334.
8. Барсболд Р. Хищные динозавры мела Монголии. Сер.: «Тр. совместной Советско-Монгольской палеонтол. экспедиции». Вып. 19. М., 1983.
9. Курзанов С. М. Авимимиды и проблема происхождения птиц. Сер.: «Тр. совместной Советско-Монгольской палеонтол. экспедиции». Вып. 31. М., 1987.











Упрощенная геохронологическая шкала мезозоя