Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Всему свое время и место
Вторичным покоем называют такое явление в жизненном цикле сезонных многолетних растений, когда растение не возобновляет рост и развитие надземных побегов с началом очередного сезона вегетации и выглядит погибшим, но через несколько лет вдруг оживает. Уникальные длительные ряды данных ежегодных наблюдений (2009–2023гг.) за локальными популяциями двух альпийских малолетних видов на постоянных площадках позволили выявить эффекты вторичного покоя в жизненном цикле этих видов, что принципиально невозможно на более коротких отрезках времени. Это открытие потребовало ревизовать данные последних лет наблюдений и провести ревизию математического формализма, который обеспечивает объективную количественную оценку меры жизнеспособности локальных популяций.
«Тише, мыши, кот на крыше!…», и мыши действительно притворяются мёртвыми, как и многие другие животные, чтобы обмануть хищника и сохранить себе жизнь. Оказывается, и растения могут применять подобную тактику выживания, только «хищником» выступают суровые условия среды и тому подобные стрессы.
Обычно в сезонном климате после окончания неблагоприятного для растений периода года (зимнего или летнего покоя) они возобновляют рост побегов и корней. Однако иногда этого не происходит, и если условия губительны для молодых побегов, то они не появляются на свет в течение всего года или на протяжении нескольких лет и растение выглядит погибшим. Но корни такого растения сохраняют жизнеспособность, и через несколько лет оно вдруг «оживает», то есть начинает снова развивать надземные побеги. В ботанической науке явление получило название вторичный покой, а в эволюционном сознании – признается как важный механизм приспособления, обеспечивающий выживание вида.
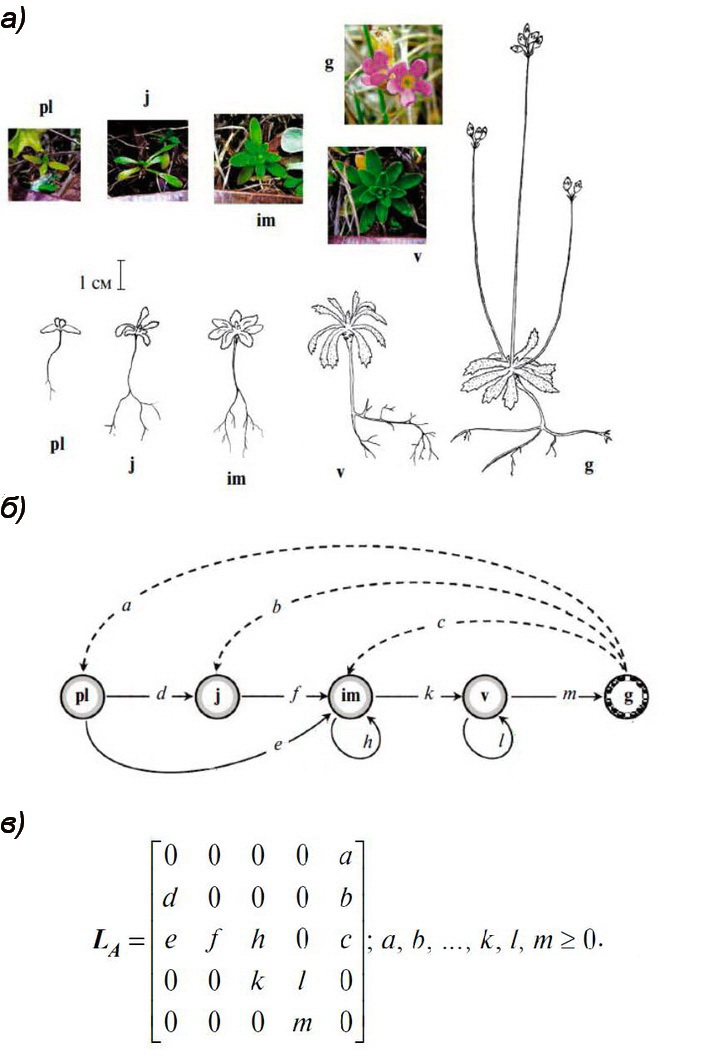
Обнаружить вторичный покой в популяциях растений весьма непросто. Нужны многолетние наблюдения («всему свое время») за фиксированными особями («… и место»), а также уверенность, что ни что иное не погубит растение за это долгое время в этом нужном месте. Поэтому структуру локальных популяций двух альпийских малолетних видов: краснокнижного проломника Androsace albana и эндемичного незабудочника кавказского Eritrichium caucasicum, – наблюдали ежегодно в августе, в течение 15 лет, на постоянных площадках, заложенных в 2009 г. на территории Тебердинского национального парка, на горе Малая Хатипара (высота над уровнем моря 2800 м). Но растения, вышедшие из состояния вторичного покоя, обнаружили только в последние 2-3 года.
Исследования проводили с целью дать объективную количественную оценку меры жизнеспособности популяций, и эта цель достигалась путём построения так называемых матричных моделей динамики популяций со стадийной структурой. Структуру фиксировали согласно известным шкалам онтогенеза (рис. 1а), а модели строили по разработанной ранее методике с помощью так называемого графа жизненного цикла (рис. 1б). В этом графе 5 вершин соответствуют пяти заявленным стадиям, а стрелки – пополнению популяции и переходам между стадиями за прошедший год, зафиксированным в наблюдениях. Годичная матрица размера 5✕5 соответствует графу согласно фундаментальному математическому правилу, и её строение, то есть расположение нулевых и положительных элементов внутри матрицы (рис. 1в), однозначно определяется этим графом. Десять положительных элементов a, b, c, …, l, m годичной матрицы – демографические параметры популяции – вычисляются однозначно по данным двух соседних лет наблюдений, давая калиброванную матрицу. Она-то и служит источником разнообразных количественных показателей, в том числе и меры жизнеспособности, данной популяции, которые вычисляются рутинным образом.
Понятно, что растения, которые ранее считались погибшими, а потом вдруг «ожили», то есть вышли из состояния вторичного покоя, требуют ретроспективной ревизии данных наблюдений, а ревизия данных – соответствующий ревизии матричного формализма: вместо состояния гибели, которое явно не присутствовало в графе жизненного цикла, нужно теперь рассматривать состояние гибели или вторичного покоя. Размерность новой годичной матрицы возрастает на единицу, и соответствующим образом модифицируется процедура ее калибровки. Процедуры калибровки, скорректированные по ревизованным данным в рамках ревизованного формализма (для обоих изучавшихся видов), изменили – хотя и в разной степени – прежнюю оценку меры жизнеспособности в сторону увеличения, подтвердив тем самым важную роль вторичного покоя как механизма адаптации к стрессовым условиям существования.
Разумеется, разные пары соседних лет наблюдений порождают годичные матрицы, которые количественно отличаются друг от друга, соответственно различаются и вычисленные по ним, годичные меры жизнеспособности. Когда же стоит задача оценить жизнеспособность популяции на основе многолетнего периода наблюдений, встаёт вопрос, как именно следует усреднить эти 14 матриц (среднее арифметическое не имеет здесь никакого смысла) или как вычислить так называемую стохастическую скорость роста при определённых сценариях случайных изменений среды. Эти задачи решались коллективом авторов до проявления эффектов вторичного покоя и будут решаться вновь в рамках ревизованных матричных формализмов.
Читайте о других исследованиях динамики популяций с помощью математических моделей авторов статьи на страницах «Журнала общей биологии»:
1. Моделирование сукцессии: можно ли заглянуть в прошлое? (к статье ЖОБ Том 83, 2022. № 5);
2. Совместная динамика популяций черники и брусники в послепожарном сосняке-зеленомошнике (к статье ЖОБ Том 81, 2020. № 4) ;
3. Черника против брусники: осредненная марковская модель сосуществования в бореальном лесу (к статье ЖОБ Том 79, 2018. № 2) ;
4. Модельный подход к определению состояния популяции растений (к статье ЖОБ Том 78, 2017. № 5);
и другие статьи.
Популярные синопсисы