Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Прямые взаимодействия клеток задают независимый ритм метаболическим процессам
В статье сделан обзор современных представлений о стабильности прямых межклеточных взаимодействий, как регулятора внутритканевых ритмов различных метаболических процессов: синтеза белка, активности протеаз и АТФ-азы, разрушения и сборки ядерной оболочки. Экспериментальные данные говорят о метаболической самосинхронизации клеток в культуре при условии постоянства среды. Представляется, что ключевым процессом межклеточных взаимодействий является активация протеинкиназ.
Основные регуляции функций высших животных осуществляет, как известно, нервная система и также отдаленные от клеточных популяций гормональные (эндокринные) влияния.
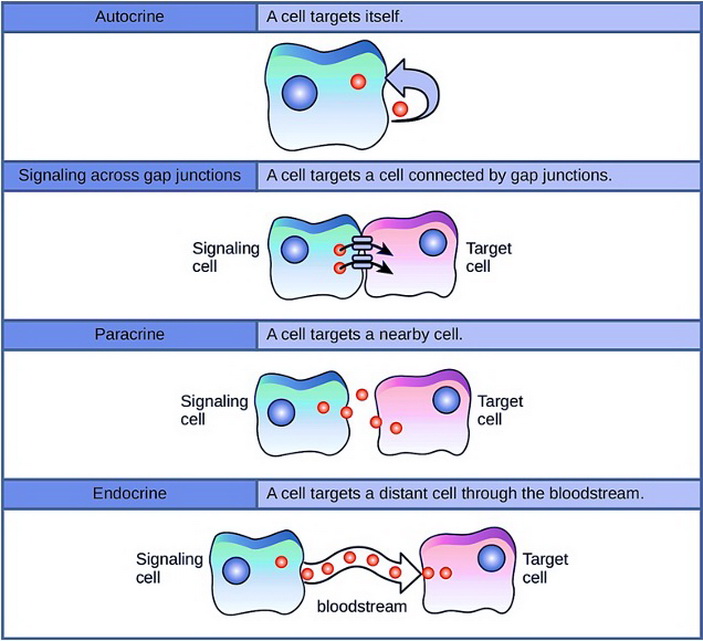
В обзоре приводятся факты прямых взаимодействий клеток (рис. 1) внутри какой-либо популяции (на примерах печени, слюнных желез, кожи), чему способствуют:
1. Перемещение из клетки в клетку ионов и малых молекул.
2. Образование специальных пограничных структур, облегчающих такой переход.
3. Сигнальные молекулы в клеточной среде, включающие процессы согласования активности клеток. Среди них найдены трансмиттеры крови: норадреналин, глутаминовая кислота, серотонин и его производное мелатонин, а также ганглиозиды.
Обоснована модель для определения и исследования прямых межклеточных отношений: плотные и редкие клеточные культуры. Индикатором таких отношений взяты околочасовые (ультрадианные) клеточные ритмы. В обзоре перечислены такие ритмы, найденные в конце 1950-х для массы клеток, а затем для синтеза белка, активности ферментов, аденилатов, включая АТФ и цАМФ, pH цитоплазмы, дыхания клеток. Ритмы выявлены у бактерий и многоклеточных, в том числе млекопитающих. В суточных плотных культурах околочасовой ритм определяется через несколько минут после начала анализа. В редких культурах ритма в это время нет.
Добавление в среду редких культур какого-либо сигнального фактора, например, фенилэфрина (фармакологический аналог норадреналина) или мелатонина, выявляет ритм в редких культурах через несколько минут. Опыт, в котором после денервации печени в ней сохраняется ритм синтеза белка, говорит о том, что в организме действуют именно факторы среды.
О стабильности прямых межклеточных взаимодействий говорит и их биохимический механизм. Использованием стимуляторов и ингибиторов процессов в экспериментах было показано, что после действия сигнала на мембрану в клетке увеличивается содержание ионов кальция в результате их выделения из цитоплазматических депо и активируются кальций-зависимые протеинкиназы. Альтернативный путь, когда сигнал активирует аденилатциклазу и в результате синтезируются АТФ и цАМФ, что приводит к активации цАМФ-зависимых протеинкиназ. В свою очередь показано, что активация протеинкиназ вызывает сдвиг в ритме синтеза белка.
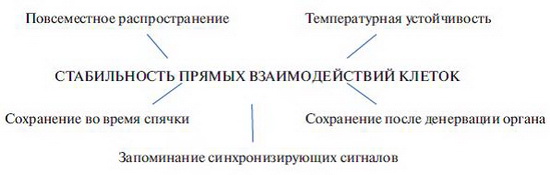
Все эти данные, изложенные в обзоре, утверждают стабильность прямых межклеточных отношений и их индикатора околочасового ритма (рис. 2). О том же говорит и температурная их устойчивость, отмеченная сначала для делений дробления, а затем для разных состояний клеток в культуре и в организме. Так, околочасовые ритмы сохраняются во время спячки животных (сусликов). Сохраняются метаболические околочасовые ритмы и при торможении ритма митозов, о чем свидетельствуют опыты с остановкой делений дробления.
В обзоре приведены наши экспериментальные данные, подтверждающие математическую модель И.М. Гельфанда и М.Л. Цетлина о самосинхронизации осцилляторов в общей среде. Получены культуры гепатоцитов с противофазными колебаниями околочасового ритма синтеза белка. В культурах из смеси равных количеств таких клеток обнаружен ритм, а не линейная кинетика.
Факты, изложенные выше, поддерживают представление о гомеодинамике, как форме гомеостаза.
Популярные синопсисы