Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Обоняние безнервных животных могло заложить основы глутаматной передачи в нервной системе

Сценарий эволюционного происхождения нервной системы, каким бы он не оказался, должен включать в себя предположения о происхождении разнообразия нейротрансмиттеров. Животные из типа Placozoa - прекрасный модельный объект для изучения этого вопроса, так как имеют одновременно сложное поведение и гены рецепторов важнейших семейств нейротрансмиттеров, а нервной системы не имеют. Мы произвели поиск, аннотацию и филогенетический анализ генов, кодирующих рецепторы глутамата, глицина и гамма-аминомасляной кислоты, в геномах двух видов Placozoa: Trichoplax adhaerens и Hoilungia hongkongensis. По скорости и обилию делеций и дупликаций гены глутаматных и ГАМК рецепторов Placozoa похожи на гены обонятельных рецепторов Bilateria. Возможно, аминокислотные нейротрансмиттеры исходно были важными химическими сигналами из внешней среды, на которые реагировала система хеморецепции древних донервных животных.
Появление нервной системы является одним из важнейших эволюционных событий на Земле. Но каким образом появилась нервная система? Этот вопрос требует дальнейшего изучения. В частности, остается неясным происхождение и многообразие нейтротрансмиттеров, включая глутамата и гамма-аминомасляной кислоты (ГАМК). Интересным объектом для изучения этого вопроса являются животные типа Placozoa: они подвижны, имеют пищевое, поисковое и даже социальное поведение, и все это при полном отсутствии нервной системы (Заглавная иллюстрация). В геноме наиболее изученного вида Placozoa, Trichoplax adhaerens, обнаружены ионные каналы, которые участвуют в передаче нервных импульсов у животных. Неизвестно, является ли отсутствие нервной системы (НС) у Placozoa первичным или вторичным. В любом случае, изучение их рецепторов даст важную информацию либо об исходных функциях у донервных животных, либо о путях смены функций бывших компонентов НС. Поэтому мы произвели поиск, аннотацию и филогенетический анализ генов, кодирующих рецепторы глутамата, глицина и ГАМК, в геномах двух видов Placozoa: Trichoplax adhaerens и Hoilungia hongkongensis.
У позвоночных животных в глутаматной передаче участвуют возбуждающие ионотропные глутаматные рецепторы (iGluR) и метаботропные глутаматные рецепторы (mGluR). В ГАМК-зависимой передаче участвуют тормозные ионотропные ГАМК-A рецепторы (GABA-A) и метаботропные ГАМК-B рецепторы (GABA-B). В восприятии глицина у позвоночных основную роль играют тормозные ионотропные глициновые рецепторы (GlyR). В геноме Trichoplax обнаружены гены для большинства рецепторов из этих семейств рецепторов, за исключением ГАМК-A и глициновых. В то же время, у представителя другого рода пластинчатых, Hoilungia hongkongensis, только 5 iGluR (рис. 1). Мы обнаружили у Placozoa большое разнообразие рецепторов iGluR, mGluR и GABA-B. Количество iGluR Trichoplax (14) сопоставимо с таковым у животных с нервной системой (17 у человека), а количество метаботропных рецепторов mGluR и GABA-B даже выше, чем у большинства Bilateria.
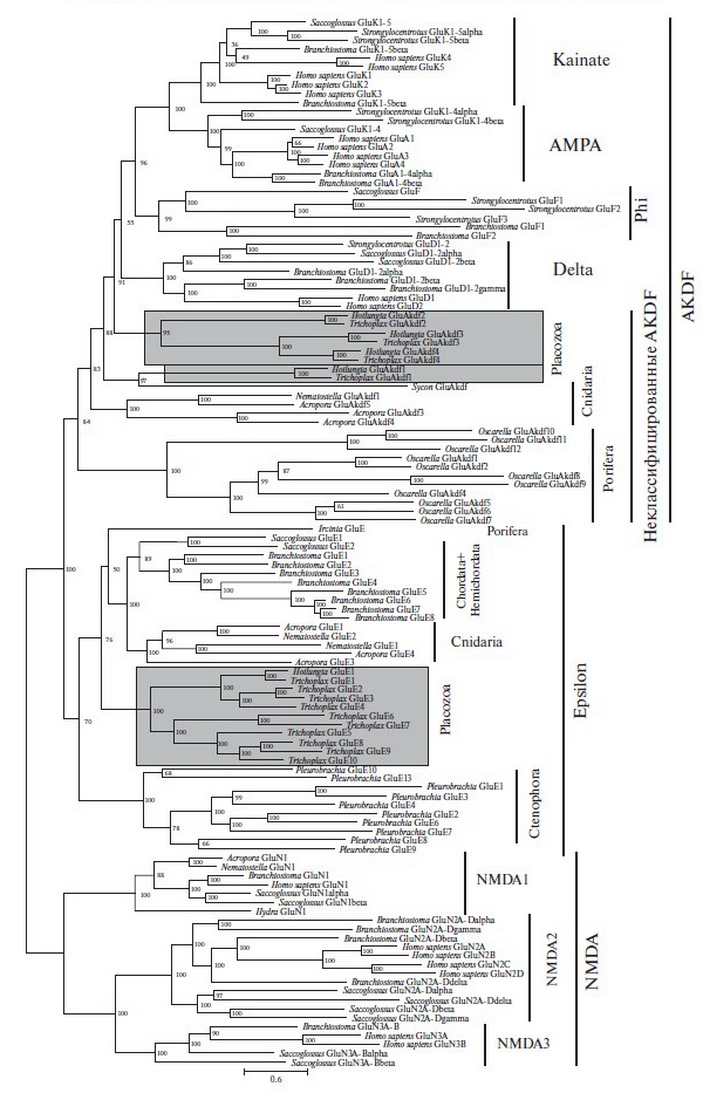
Сравнение аминокислот, участвующих в связывании лигандов, указывает на большое разнообразие лигандной специфичности iGluR Trichoplax. Функции iGluR Placozoa не могут ограничиваться глутаматной и глициновой передачей, они также участвуют в восприятии других веществ, причем скорее из внешней среды. Гены рецепторов iGluR, mGluR и ГАМК-В Placozoa неожиданно склонны к дупликациям и делециям, причем в эволюции рода Hoilungia преобладала потеря ионотропных и дупликация метаботропных рецепторов. Рецепторы Placozoa по эволюционной динамичности сходны с обонятельными рецепторами Bilateria. Поэтому мы можем ожидать, что большинство mGluR-подобных и ГАМК-В-подобных рецепторов Placozoa участвуют в хеморецепции внешних стимулов.
Чтобы вещество выполняло функции трансмиттера, необходима слаженная работа систем синтеза и рецепции этого вещества. Одновременное возникновение обеих систем эволюционно крайне маловероятное событие. Поэтому для появления трансмиттеров, по-видимому, необходимо, чтобы хотя бы одна из биохимических систем возникла путем смены функции. Следовательно, возможны два эволюционных сценария. Первый: имеется система рецепции вещества, но само вещество первоначально поступает в организм из внешней среды, а затем сигнал интернализуется. Второй: существует система синтеза вещества, но оно выполняет например, защитные, а не сигнальные функции. Затем возникают рецепторы, и вещество становится трансмиттером. Первый сценарий показан для тироидных гормонов животных, происходящих от вторичных метаболитов водорослей, а второй более вероятен для нейропептидов и оксида азота (NO). Результаты нашего исследования указывают на то, что хеморецепторная система была важным источником разнообразных рецепторов для возникающей нервной системы, а будущие аминокислоты-нейротрансмиттеры (глутамат, ГАМК, глицин) были релевантными внешними сигналами для первых животных, еще не имевших нервной системы.

Популярные синопсисы