Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Теория и практика: экологические классификации видов растений

Статья содержит обзор экологических классификаций видов растений и различий характеристик, определяющих группировки (типы) растений в зависимости от выбранного подхода. Можно выделить два основных способа систематизации знаний в экологических исследованиях сообществ растений: определение групп по внешнему облику растений или по отношению видов к экологическим факторам. Авторы предполагают, что оперирование функциональными группами растений, в отличие от использования систем жизненных форм, более эффективно в геоботанических исследованиях.
Классификации разного рода систематизируют большой объем данных, помогают схематизировать эволюцию и разнообразие биологических объектов. Разнообразие растительных сообществ также подвергается анализу и классификации (см. синопсис и статьи в ЖОБ «Основные тенденции развития теории синтаксономии и динамики растительных сообществ», «Концепция растительного сообщества: история и современное состояние»). Для классификации сообществ необходимо понимание экологических характеристик видов, составляющих разные типы сообществ, выявление закономерностей видовой мозаики в сходных и различающихся сообществах. В статье авторы предлагают обзор экологических классификаций видов растений и описание важных характеристик, которые определяют группировки растений согласно разным классификациям.
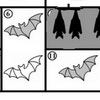
Имеется два подхода к экологической классификации видов растений, по аналогии с классификацией сообществ: физиогномический и флористический. Физиогномические экологические классификации основаны на выделение групп видов по сходству внешнего облика растений, флористические – по сходству требований к экологическим условиям. В первом случае принято различать жизненные формы растений (ЖФ) и формы роста (ФР). Считается, что ЖФ, т.е. собственно морфология растения, является следствием адаптации растения к условиям среды, тогда как ФР определяется «внутренним механизмом» и относительно независима от комплекса внешних условий. Надо отметить, что в отечественных исследованиях понятие ФР непопулярно, зато применение нашли несколько систем ЖФ. Фундамент деления растений по ЖФ заложил философ Теофраст за 300 лет до н.э. Он выделил три типа ЖФ – деревья, кустарники и травы. Эти три типа до сих пор применяются в качестве упрощенной схемы для разного рода обобщающих исследований (см., например, статью в текущем номере ЖОБ «Современная и мел-кайнозойская диверсификация покрытосеменных»). Однако эта система получила развитие в работах ботаников, которое пошло по пути увеличения числа типов и подтипов ЖФ. Так, например, в системе Гумбольдта (1806) насчитывается 16 типов форм, среди которых пальмы, кактусы, банан, а в системе Друде (1896) число подтипов достигло 60, объединенных в 7 основных типов: древесные растения, суккуленты, лианы, эпифиты, злаки, лишайники, мхи. Одной из наиболее широко известных и применяемых физиогномических классификаций является система К. Раункиера. Он предложил систему основанную всего на одном признаке – положении и способе защиты почек возобновления в течение неблагоприятного периода (холодного или сухого) (рис. 1).

1 – фанерофиты (тополь); 2 – хамефиты (черника); 3 – гемикриптофиты (одуванчик); 4 – геофиты (тюльпан); 5 – терофиты (фасоль). Зимующие части находятся под пунктирной линией. Фото с сайта eco-rasteniya.ru
Понятие ЖФ может быть акцентировано применением терминов функциональные типы растений или функциональные группы растений, в выделении которых особое значение приобретают такие параметры как продуктивность, индекс листовой поверхности, сезонная ритмика и проч. Эффективность системы ЖФ Раункиера была показана Р. Уиттекером (1980) при сравнении флористического состава биомов. Он показал, например, что во влажных тропических лесах преобладают фанерофиты, в лесах умеренно холодной зоны, тундре и степи – гемикриптофиты, в полупустыне – хамефиты, а в пустыне – терофиты.
Система Раункиера получила дальнейшее развитие в работах многих исследователей. Хорошо известна работа Мюллер-Дембуа и Элленберга (1974) , которые использовали для классификации дополнительные признаки: тип питания, анатомическое строение, сезонная ритмика, степень одревеснения тканей и многие другие. Хотя система Мюллер-Дембуа и Элленберга логически и экологически обоснована, использовать на практике эту систему затруднительно, поскольку такая подробная детализация слишком сложна для рационального анализа сообществ. Сходной сложностью отличается и система И.Г. Серебрякова, поэтому отечественные исследователи предпочитают использовать только некоторые основные ранги этой системы.
Я. Баркман (Barkman, 1988) развил классификацию ФР, которая была противопоставлена ЖФ. В ней деревья делятся исключительно по морфологическим признакам наименее связанным с условиями обитания (насколько это вообще возможно, конечно). На первом уровне растения делятся на свободно плавающие, прикрепленные и имеющие корни. Понятно, что в одной системе рассматриваются разнообразные по систематическому положению виды – и водоросли, и сосудистые растения, и мхи. На следующем уровне виды делятся на типы по характерным представителям и т.д. Система отличается тем, что на всех уровнях есть строгие (формализованные) правила выделения групп растений.
Другим путем пошли исследователи, которые попытались сделать систему ЖФ более пригодной для применения на практике. Предполагалось перейти от классификации формы к функционированию растения. Система функциональных типов растений (ФТР) основана на выделении типов, различающихся по функциональным признакам. Такие типы выступают в роли доминантов биомов и отвечают за их основные функциональные характеристики. Например, ФТР используются в первую очередь при аэрокосмической съемке. Выделенные на основе ФТР биомы различаются по таким существенным характеристикам, как индекс листовой поверхности, первичная биологическая продукция и показатель испарения влаги.
Отдельная методика используется для выделения функциональных групп растений (ФГР). Как правило, ФГР отражают поведение растений в ходе сукцессии, например, при пирогенной сукцессии или пастбищной дигрессии.
Методика выделения экологических групп существенно отличается от таковой по морфологическим признакам (даже учитывая связь морфологии с внешними условиями). Основные подходы к выделению экологических групп видов сформированы под воздействием концепции непрерывности Л.Г. Раменского. Дискретность биомов признается условной, распределение видов растений в пространстве определяется градиентами различных экологических факторов. Градиент определенного фактора разбивается (условно) на конечное число градаций. Предполагается что, экологические группы составляют виды, у которых модальные классы распределения по данному градиенту лежат в рамках установленных градаций. Понятно, что группы, выделенные по градиенту одного фактора, будут распадаться (в той или иной степени) при рассмотрении распределений видов относительно другого фактора. Поэтому для выделения таких групп не используется более 2-3 наиболее «говорящих» факторов. Фитоценологи часто выделяют экологические группы по отношению к ведущим факторам среды, таким как увлажнение (мезофиты, гигрофиты, ксерофиты и т.п.), засолению (галофиты, гликофиты, и т.п.). Широко используется градиентный анализ вдоль одного фактора среды: экоклины (влажность), топоклины (высота над уровнем моря), хроноклины (в случае сукцессии).
В рамках флористического подхода следует упомянуть распространённый в отечественной геоботанической литературе метод выделения эколого-ценотических групп (ЭЦГ). Выделение групп происходит на основе видов-индикаторов типов сообществ, определяемых в ходе флористического описания сообществ по методу Браун-Бланке. При определении видов-индикаторов применяют как экспертную оценку, так и статистическую оценку значимости определения.
Авторы предполагают, что оперирование функциональными группами растений, в отличие от использования систем жизненных форм, более эффективно в геоботанических исследованиях.
С 2018г. полные тексты статей «Журнала общей биологии» за последнее десятилетие доступны авторизованным пользователям на ресурсе Научной электронной библиотеки eLIBRARY.RU
Популярные синопсисы