Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Лектины заменяют растениям иммунную систему
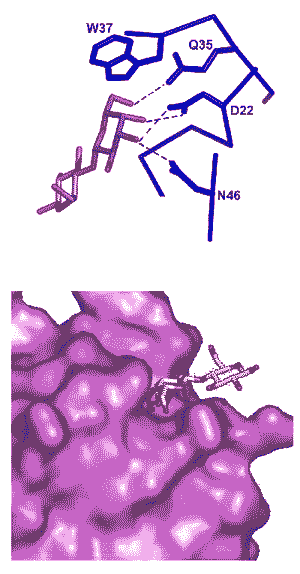
Лектины – белки, способные узнавать и избирательно связывать разнообразные углеводы – помогают растениям бороться с патогенными бактериями, грибами и вирусами. Кроме того, они регулируют взаимоотношения с симбиотическими бактериями. Хотя набор лектинов ограничен, они обеспечивают точное прочтение «углеводного кода» микробов благодаря дополнительным «хитростям», таким как скоординированная работа нескольких разных лектинов и распознавание углеводов, характерных для больших групп потенциальных патогенов.
На нашем сайте ранее был опубликован популярный синопсис статьи Ф. М. Шакировой и М. В. Безруковой «Современные представления о предполагаемых функциях лектинов растений», где вкратце рассказано о лектинах и их свойствах. Статья Александра Валентиновича Бабоши из Главного ботанического сада РАН продолжает эту тему. В статье делается акцент на тех функциях растительных лектинов, которые можно назвать «иммунологическими». Речь идет прежде всего о распознавании и обезвреживании патогенных вирусов, грибов и бактерий. Лектины выполняют у растений, по сути дела, ту же функцию, что и иммуноглобулиновые белки у животных. Конечно, нужно помнить, что по своей природе лектины не имеют ничего общего с иммуноглобулинами (не считая того, что те и другие являются белками). Иммуноглобулины распознают «непрошенных гостей» (например, бактерий) чаще всего по их белкам, а лектины – по углеводам.
«Иммунологические» функцие лектинов довольно разнообразны. Многие лектины растений подавляют рост паразитических грибов. Эти лектины распознают молекулы хитина и других углеводов в стенках грибных клеток, и прикрепляются к ним, мешая грибу расти. Это происходит обычно в концевых участках грибных нитей – там, где гриб растет. В зрелых (давно сформировавшихся) участках оболочек грибных клеток молекулы хитина защищены от взаимодействия с лектинами, но в молодых участках лектинам удается «ухватиться» за хитин и заторомозить рост гриба.
Лектины также выполняют роль рецепторов, реагирующих на специфические углеводы клеточных оболочек патогенных микроорганизмов. По-видимому, многие слабо изученные растительные лектины, присутствующие в клетках в небольших количествах, выполняют функции рецепторов. Иногда бывает и наоборот: лектины, имеющиеся в оболочке патогена, могут регистрироваться растительными рецепторными комплексами, в состав которых входят молекулы углеводов. Иными словами, лектин может быть как рецептором (в этом случае лигандом – распознаваемым веществом – является углевод), так и лигандом (в этом случае в роли рецептора выступает углевод, или, точнее, молекулярный комплекс, в состав которого входит углевод).
Самый интригующий вопрос, связанный с иммунологическими функциями лектинов растений, состоит в следующем. Число различных патогенных бактерий, вирусов и грибов необозримо велико. К тому же все эти паразиты быстро эволюционируют, так что растениям часто приходится сталкиваться с новыми видами и штаммами патогенов. Между тем врага надо вовремя распознать, чтобы растение успело включить защитные механизмы (например, начать синтез растительных антибиотиков – фитоалексинов). При этом у растений нет возможности «изобретать» новые защитные белки по мере надобности путем внесения изменений в геном своих клеток, как это делают позвоночные животные (см.: Мутагенез в лимфоцитах — результат целенаправленного изменения ДНК и последующей «неточной починки», «Элементы», 3.09.07). По крайней мере, до сих пор подобные механизмы у растений не обнаружены. Единственный эффективный инструмент для узнавания непрошенных гостей, который есть у растений – это лектины. Но набор лектинов ограничен. Как же с их помощью растения идентифицируют миллионы разных врагов?
В поисках ответа на этот вопрос А.В.Бабоша рассматривает по отдельности две области применения растительных лектинов: распознавание «своих» и распознавание «чужих».
Распознавание «своих». Лектины играют ключевую роль в установлении симбиотических отношений с клубеньковыми бактериями (см. ссылки внизу). В этом случае очень важно не ошибиться, не спутать «свою» бактерию с чужой. Точность идентификации при этом обеспечивается слаженной работой многих разных лектинов, которые, по-видимому, вместе осуществляют всесторонний анализ «углеводного кода» бактерии. Кроме того, процесc опознания является двусторонним: у симбиотических бактерий есть свои лектины, при помощи которых они «ощупывают» поверхность корешков своего растительного партнера и проверяют его «углеводный код».
Роль лектинов в симбиозе с клубеньковыми бактериями была подверждена генно-инженерными экспериментами: клевер, которому пересадили ген лектина гороха, стал вступать в симбиоз с клубеньковой бактерией, с которой клевер в норме в симбиоз не вступает. Лядвенец рогатый (Lotus corniculatus), которому пересадили ген одного из лектинов сои, начал вступать в симбиоз с бактерией – симбионтом сои, хотя до формирования полноценного симбиоза дело в данном случае не дошло. Эти и другие эксперименты не только доказали роль лектинов в распознавании бактериального симбионта, но и показали, что процесс опознания является сложным и многоступенчатым.
Роль лектинов в формировании симбиотических образований (клубеньков) не ограничивается распознаванием симбионта. Некоторые растительные лектины стимулируют рост клубеньковых бактерий. При этом их действие может быть очень избирательным: например, лектин сои активизирует бактерию Bradyrhizobium japonicum (симбионта сои), однако лектины других растений, даже очень похожие по структуре на соевый лектин, не действуют на эту бактерию. Предполагается, что растительные лектины на самом деле оказывают на бактерий неблагоприятное воздействие. Дело в том, что если клубеньковые бактерии находятся в благоприятных условиях, они не вступают в симбиоз с корнями растений, предпочитая жить «на свободе». К вступлению в симбиоз их побуждает, например, недостаток питания. Возможно, растение не «инфицируется» клубеньковыми бактериями, а «берет их в плен», причем лектины служат средством агрессии. Если это так, то становится легче понять, как лектины, исходно служившие для борьбы с бактериями, стали использоваться для установления симбиотических отношений с ними. Поистине, от вражды до дружбы – один шаг.
Кроме того, лектины участвуют в подавлении защитного ответа растения на «инфицирование» клубеньковыми бактериями. Это необходимо для развития симбиоза, но довольно опасно, поскольку создает брешь в защитной системе растения. Возможно, именно поэтому симбиозы растений с клубеньковыми бактериями столь избирательны (каждое растение может вступать в симбиоз лишь с очень узким кругом бактерий – потенциальных симбионтов).
Распознавание «чужих». Чтобы распознавать широкий круг патогенов при помощи небольшого набора лектинов, растения должны использовать какие-то «обобщающие» механизмы. Иными словами, защитные лектины должны реагировать на углеводы, очень широко распространенные среди потенциальных патогенов. Например, на хитин, входящий в состав клеточной стенки большинства грибов (см. выше). Однако известны и случаи избирательного взаимодействия. Например, часто бывает, что одни штаммы какого-нибудь вида бактерии эффективно обезвреживаются растительными лектинами, а другие – нет. Оказалось, что некоторые бактерии научились выделять в больших количествах во внешнюю среду специальные полисахариды, которые связываются с защитными растительными лектинами и не дают им добраться до самой бактерии.
В целом механизмы «лектинового распознавания» патогенов менее сложны и нет так специфичны, как взаимодействие лектинов с симбиотическими бактериями. Это и понятно: в случае формирования симбиоза в ходе эволюции у обоих партнеров есть больше времени, чтобы тонко «подогнать» друг к другу молекулярные системы взаимного узнавания. Борьба с патогенными микроорганизмами требует от растения гораздо большей оперативности, что способствует выработке неспецифических средств защиты с широким спектром действия. Это подверждается опытом селекционеров, выводящих сорта растений, устойчивых к какому-нибудь патогену. Если механизм устойчивости специфичен (растение безошибочно распознает именно данный вид патогена), то быстро эволюционирующий патоген обычно легко «уходит» от этой специфической системы узнавания, и только постоянные усилия больших коллективов селекционеров позволяют растению не проиграть в этой гонке. Ясно, что в естественных условиях растениям лучше полагаться на «обобщающие» механизмы распознавания возможных патогенов, хотя при этом, очевидно, снижается эффективность защитных систем (например, один и тот же защитный ответ будет включаться при контакте и с очень опасными, и с безвредными бактериями).
Лектины в иммунитете животных. Большинство беспозвоночных животных тоже не имеют такой эффективной системы выработки новых защитных белков, как у позвоночных. Поэтому им, как и растениям, для защиты от микробов могут очень пригодиться лектины. Это предположение подтверждается множеством фактов. Впрочем, и у позвоночных лектины играют важную роль в неспецифическом (врожденном) иммунитете.
Некоторые патогены научились использовать защитные лектины животных к собственной выгоде. Например, многие вирусы проникают в клетки человека, прицепившись к молекулам лектинов, расположенным на поверхности клетки. Поэтому выработка защитных лектинов животными может быть «палкой о двух концах». Например, высокий уровень одного из лектинов (маннозо-специфичного лектина) в сыворотке крови человека способствует устойчивости к ряду бактериальных и вирусных заболеваний, включая ВИЧ, однако повышает восприимчивость к некоторым другим болезням, таким как туберкулез и лепра.
Лектины растений, введенные в организм животного, могут оказывать действие, сходное с действием собственных лектинов животного. Например, они могут влиять на работу различных компонентов иммунной системы. Они могут даже стимулировать гибель (апоптоз) раковых клеток. Один из лектинов омелы (Viscum album) широко применяется в медицине для лечения рака.
Автор заключает, что лектины, взаимодействующие с поверхностными углеводами патогена, представляют собой некий универсальных механизм, позволяющий воздействовать на рост микроорганизмов. Этот механизм используется как растениями, так и животными.
Гликокод. Некоторые исследователи отмечают, что количество информации, содержащейся в углеводах (например, в полисахаридах клеточной стенки) значительно больше, чем это необходимо для выполнения такими полисахаридами чисто механических функций. Углеводы – очень подходящая материальная основа для кодирования информации, в некотором смысле даже более подходящая, чем белки и нуклеиновые кислоты. Например, две одинаковые аминокислоты можно соединить друг с другом только одним способом (из них получается только один дипептид). То же и с двумя одинаковыми нуклеотидами – из них более одного динуклеотида никак не получится. Однако из двух молекул глюкозы, соединяя их друг с другом по-разному, можно получить целых девять разных дигликозидов. Углеводы, в отличие от белков и нуклеиновых кислот, могут образовывать не только линейные полимеры, но и ветвящиеся. Вполне возможно, что роль углеводов в кодировании биологически значимой информации (и в системах взаимного распознавания и идентификации) сильно недооценивается. Биологическую информацию, записанную языком углеводных остатков, предложено называть «гликокодом». Ключевую роль в прочтении и интерпретации гликокода играют, разумеется, лектины.
Казалось бы, этому противоречат имеющиеся представления о низкой эффективности и специфичности (избирательности) лектинов как распознающих молекул. В большинстве проведенных экспериментов эффективность лектинов в этом плане оказывалась на несколько порядков ниже, чем у иммуноглобулинов. Однако, как указывает А.В.Бабоша, в таких экспериментах обычно в качестве лигандов (распознаваемых и связываемых веществ) используются моносахариды, то есть самые простые из углеводов. Между тем ряд фактов указывает на то, что многие лектины на самом деле специализируются на связывании куда более сложных углеводов (с которыми проводить эксперименты гораздо менее удобно). Поэтому очень может быть, что эффективность и избирательность лектинов сильно недооценивается.
См. также о лектинах:
Лектины – белки, специализирующиеся на распознавании углеводов (по статье: Ф. М. Шакирова, М. В. Безрукова. Современные представления о предполагаемых функциях лектинов растений).
О симбиозе растений с азотфиксирующими (клубеньковыми) бактериями:
Изменение гена, необходимого для симбиоза растений с грибами, привело к формированию симбиоза с азотфиксирующими бактериями, «Элементы», 12.03.08.
От биохимического сотрудничества – к общему геному (по статье: Н. А. Проворов, Е. А. Долгих
Метаболическая интеграция организмов в системах симбиоза).
Популярные синопсисы