Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Специализация открывает дорогу прогрессу

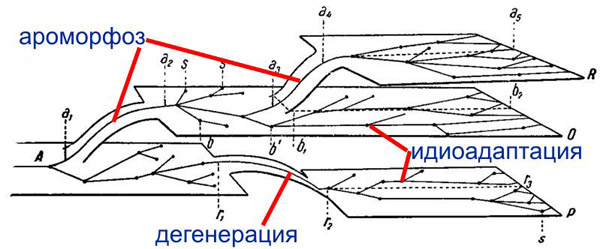
По мнению зав. кафедрой биологической эволюции биофака МГУ А.С.Северцова, важнейшим критерием ароморфозов – прогрессивных эволюционных изменений, связанных с усложнением организации – является расширение условий обитания (адаптивной зоны) организмов. Вопреки расхожему мнению о происхождении новых групп организмов преимущественно от неспециализированных представителей предковой группы, ароморфозам часто предшествует специализация в узкой пограничной области адаптивной зоны предков.
Понятие «ароморфоз» было введено А.Н.Северцовым в 1914 для того, чтобы формализовать и обобщить представления о тех эволюционных изменениях, которые в просторечии обычно именуют «прогрессивными» или просто «самыми важными».
А.Н.Северцов считал главными критериями ароморфоза усложнение организации и интенсификацию обмена веществ (повышение энергии жизнедеятельности). Концепция ароморфоза оказалась весьма полезной и востребованной. Последователи и ученики А.Н.Северцова, в том числе С.А.Северцов и И.И.Шмальгаузен, продолжали развивать учение об ароморфозе. Они придавали большое значение еще одному критерию ароморфоза, который можно условно назвать «экологическим критерием». Ароморфоз, по их мнению, это приспособление широкого назначения, позволяющее организмам существовать в более сложных и разнообразных условиях среды, чем это было свойственно их предкам.
В статье профессора А.С.Северцова, заведующего кафедрой биологической эволюции биофака МГУ, дается обзор вглядов разных исследователей и излагается авторское понимание ароморфоза.
А.С.Северцов подвергает критике понятие «ключевого ароморфоза» Н.Н.Иорданского, отмечая, что «в сложном комплексе преобразований организации, характеризующем ароморфную эволюцию, едва ли можно выделить единственное адаптивное изменение, служащее пусковым механизмом для всех других адаптивных изменений». Автор также подчеркивает, что нельзя оценивать ароморфность по таксономическому уровню (например, оболочники – это подтип, возникший на основе общей дегенрации, то есть упрощения организации).
По мнению А.С.Северцова, ароморфозом следует считать «такой комплекс преобразований организации, который позволяет потомкам расширить адаптивную зону, свойственную их предкам».
При ароморфозе происходит не смена адаптивных зон, а добавление вновь осваиваемой среды обитания к той, которая унаследована от предков. Например, у предков четвероногих – кистеперых рыб отряда Rhipidistia – адаптивная зона была довольно узкой. Эти рыбы жили в мелких, бедных кислородом водоемах, и были подстерегающими хищниками, охотившимися на других рыб. Древнейшие четвероногие сохранили ту же среду обитания и тот же способ питания, но вдобавок приобрели способность передвигаться вне воды. Таким образом, они не сменили свою адаптивную зону, а расширили ее.
По мнению А.С.Северцова, приобретение птицами теплокровности – это ароморфоз, потому что данный признак расширил их условия обитания по сравнению с предками – рептилиями; он адаптивен (полезен) при любом способе существования птиц в их расширившейся адаптивной зоне. Однако этого нельзя сказать о таком эволюционном событии, как превращение передних конечностей в крылья. Этот признак полезен только при некоторых образах жизни, т.е. лишь в некоторых областях адаптивной зоны. При возврате птиц к наземному образу жизни крылья у них редуцируются за ненадобностью (в отличие от теплокровности).
А.С.Северцов отмечает, что многие ароморфозы в ходе становления четвероногих первоначально имели значение частных приспособлений в исходной для наземных позвоночных адаптивной зоне кистеперых рыб – рипидистий. Эти приспособления во многих случаях развивались весьма медленно и постепенно, что противоречит точке зрения о резком ускорении ароморфной эволюции в процессе расширения адаптивной зоны.
Вопреки известному «правилу неспециализированного предка», ароморфные таксоны часто происходят как раз от высокоспециализированных предков. Это было показано рядом авторов (М.С.Гиляровым, Л.П.Татариновым, Э.И.Воробьевой, А.С.Северцовым) на материале происхождения насекомых, млекопитающих, тетрапод. По мнению А.С.Северцова, специализация «в узкой, пограничной адаптивной зоне, граничащей с той, которую затем осваивает ароморфный таксон» является важнейшей предпосылкой ароморфных эволюционных преобразований.
Так, весь процесс эволюции рипидистий – это процесс специализации, как морфологической, так и экологической. Разделение костных рыб на лопастеперых (двоякодышащие + кистеперые) и лучеперых произошло в начале девона, причем лучеперые заняли в общем более глубоководные местообитания, а лопастеперые оказались приуроченными в основном к мелководьям. Затем лопастеперые разделились на двоякодышащих и кистеперых, что вело, по мнению А.С.Северцова, к сужению кормовой базы рипидистий. Кистеперые (рипидистии) оставались хищниками в мелких водоемах. В этом автор видит экологическую специализацию. Что касается морфологической специализации, то она проявилась «в развитии гулярного механизма вентиляции жабр; подразделении осевого черепа на подвижные относительно друг друга этмосфеноидный и отико-окципитальный блоки; в прогрессивном развитии мясистых парных плавников, обеспечивающих опору на дно и в ряде адаптаций подъязычно-жаберного аппарата и зубов (Воробьева, 1992; А.С.Северцов, 1990)».
В заключительной части статьи А.С.Северцов обсуждает явление многочисленных асинхронных параллелизмов, наблюдающихся как в ходе ароморфной эволюции («маммализация» - становление млекопитающих, «артроподизация» - становление членистоногих, и т.д.), так и при иных (неароморфных) эволюционных преобразованиях. Читатели могут познакомиться с подборками материалов и литературных источников по данной проблеме в наших обзорах «Ароморфозы и параллельная эволюция»; «Параллелизмы и гомологическая изменчивость: причины и следствия».
Популярные синопсисы