«Сотворение Земли». Главы из книги

Глава 9. Великое кислородное событие, акт первый: литосфера
Благодаря оттоку водорода с малого небесного тела в космос Марс и превратился в Красную планету: его цветовая гамма обусловлена окислением железосодержащих минералов, но свободного кислорода там не осталось.
На Земле насыщение кислородом атмосферы и верхних слоев гидросферы, с которыми воздух активно перемешивается вследствие штормов и ливней, обычно рисуется как поступательный процесс с резким скачком уровня этого газа примерно до 2% и затем почти до нынешних объемов кислорода в воздушной оболочке Земли (20,95%). Более тщательные исследования минерального, элементного и изотопного состава протерозойских вод (конечно, в виде выпавших из них осадков) показывают, что газ накапливался очень медленно. Даже в самом конце протерозойского эона уровень кислорода был неустойчив и не выходил за пределы 0,02%. Точка невозврата, 1–2%, вероятно, была преодолена лишь на рубеже протерозоя и палеозоя (в эдиакарском-кембрийском периодах, 635–485 млн лет назад).

Рис. 4.1, ж. Относительная доля в атмосфере кислорода, метана и углекислого газа в архейском и протерозойском эонах (3,5–0,5 млрд лет назад)
И пусть уровень кислорода в начале протерозойского эона достиг всего 1–2%, этот рубеж полностью заслуживает своего научного признания как Великое кислородное событие, поскольку в архейской атмосфере живительного газа было не более 0,0001% (рис. 4.1, ж). С этим событием связаны существенные изменения в минералогическом составе горных пород (число минералов более чем удвоилось: с 1500 до 4000), наступление ледниковых эпох, пожалуй, самых суровых в истории планеты, и появление, а затем и бурная диверсификация (рост разнообразия) эукариот (рис. 4.1, к, м).
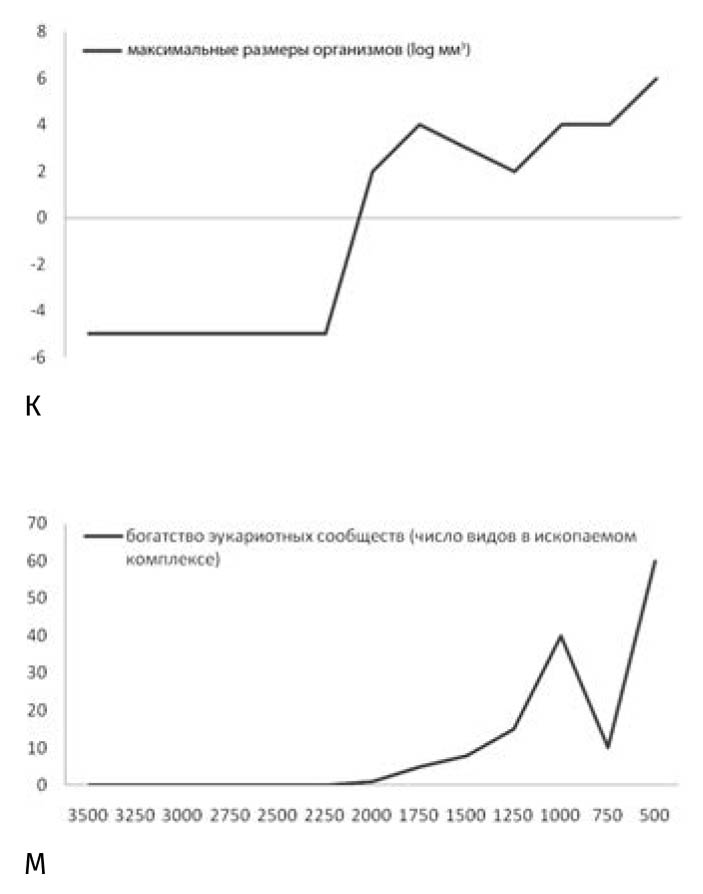
Рис. 4.1, к, м. Максимальный размер организмов (к) и богатство эукариотных сообществ (м) в архейском и протерозойском эонах (3,5–0,5 млрд лет назад)
Самым ярким свидетельством Великого кислородного события служат красноцветы — песчаники и конгломераты с песчинками и гальками, покрытыми водными окислами железа [FeO(OH); 2Fe2O3 × 3H2O)] — ржавчиной, которые начинают встречаться в континентальных отложениях возрастом от 2,43 млрд лет и моложе. А поскольку базальты — одна из наиболее развитых на поверхности суши пород — содержат железо в закисной форме, появление свободного кислорода вызвало окисление железа и привело к повсеместному покраснению этих мафических пород. Земля из черной планеты почти в одночасье стала красной, но не навсегда, как Марс. И не весь кислород остался связанным в минералах. Именно массовое появление новых минералов свидетельствует о повышении роли свободного кислорода во всех земных процессах. Например, его использовали серные бактерии для окисления сульфидов до сульфатов. И в ископаемой летописи 2,3 млрд лет назад появился гипс (CaSO4 × 2H2O). Хотя этот минерал весьма нестоек — быстро растворяется, заместившие его кремнезем и доломит сохранили присущую кристаллам этого сульфата форму: гипсовые розы и воронки, когда-то росшие на теплых пересыхающих мелководьях.
Возникновение обширного класса разбухающих глинистых минералов также связано с повышением уровня кислорода. Присутствие водонасыщенных глин, весьма недолговечных, выявляется по увеличению доли силикатных минералов с высоким содержанием δ18О в гранитной магме. Этот изотопный избыток является следствием усилившегося выветривания, к чему привело возрастание роли углекислого газа, и низкотемпературного диагенеза при изотопном обмене минералов с водной средой. Одновременно этот скачок изотопной подписи свидетельствует о появлении значительного резервуара обогащенных δ18О осадочных пород — глинистых сланцев и аргиллитов. (Углекислый газ, растворяясь в воде, превращается в угольную кислоту, которая изменяет многие минералы и изотопную подпись.)
Действительно, в архейских отложениях мощные глинистые толщи — главные продукты выветривания кристаллических горных пород — отсутствуют. Усиление выветривания отнюдь не абстрактный вывод, который следует из природы кислорода как сильнейшего окислителя: недаром французский химик Антуан Лавуазье назвал этот газ «кислоту рождающим» (франц. oxygène). Масштабы явления можно «поверить цифрой» как раз по росту объема глинистых минералов: темпы формирования глин резко увеличиваются в протерозое и продолжают нарастать до наступления фанерозойского эона. Все это является прямым следствием повышения уровня кислорода в атмосфере и, следовательно, побочным эффектом деятельности цианобактериальных сообществ.
Изменение окислительно-восстановительных условий океана привело к формированию новых типов руд — урановых, молибденовых, ванадиевых, а также фосфоритов и шунгитов. Например, органические черные сланцы стали накопителем урана — четверть мировых запасов руд этого радиоактивного металла образовались 2,4–2,2 млрд лет назад. В обогащенной кислородом среде U6+ выносится с суши, образует в морской воде трикарбонат уранила [UO2(CO3)34−], погружается в бескислородные глубины, восстанавливается до U4+ и оседает на отрицательно заряженных органических молекулах, образуя уранинит. Причем важнейшим механизмом восстановления и, следовательно, концентрации этого элемента является деятельность микробов, подобных железо- и сульфатвосстанавливающим бактериям.
Кислородное выветривание привело и к мобилизации фосфата, который активно использовался цианобактериями и накапливался как в отмершем органическом веществе, так и в виде карбонатапатита [Ca5(PO4,CO3)3F]. Из этого минерала на Индостанском щите даже построены строматолиты, образующие пласты мощностью 5–35 м и содержащие до 37% фосфата. Богатые фосфатом породы возрастом 2,1–1,9 млрд лет сформировались и в других частях планеты. Причем изотопная подпись углерода в этих отложениях прямо указывает, что к их появлению были причастны фототрофы.

Рис. 4.1, е. Изотопная подпись углерода как показатель продуктивности биоты в архейском и протерозойском эонах (3,5–0,5 млрд лет назад)
Наконец, кривая соотношения изотопов углерода действительно стала кривой с резкими перепадами значений этого соотношения от 0 до +13‰ в интервале 2,32–2,06 млрд лет (рис. 4.1, е). Этот скачок, названный событием Ломагунди-Ятулий (по отложениям в Зимбабве и Карелии, где изотопная подпись особенно резко сдвигается в сторону положительных значений), указывает на значительные изменения в продуктивности фотосинтезирующих организмов, что, в свою очередь, отражает рост поступления биогенных элементов (фосфор, железо) с суши под воздействием кислородного выветривания и наземной биоты. Например, сидерит, который входил в состав полосчатых железистых формаций, с частью этих отложений уже оказался на суше, и его окисление обеспечивало цианобактерии сразу и железом, и карбонатом для образования органического вещества.
Кроме того, доля элементов, которые легко растворялись в бескислородном океане, уменьшилась по отношению к тем, что переносятся более насыщенной кислородом водой. К примеру, возьмем соотношение цинка и железа в карбонатных отложениях: оба этих элемента имеют одинаковую растворимость в морской воде и поступают из одних и тех же источников (в основном гидротермальных). Однако с появлением кислорода Fe2+ начинает окисляться до Fe3+ и выводится из раствора в виде окислов, тогда как для Zn2+ ничего не меняется — ионы этого металла продолжают осаждаться в карбонатах и соотношение Zn/Fe растет. Закономерным образом колеблется и фоновое содержание урана и молибдена — элементов, очень чувствительных к окислительно-восстановительному потенциалу среды. Резкие скачки в соотношении различных элементов и изотопов на архейско-протерозойской границе указывают на смену окислительно-восстановительной обстановки в древнем океане (рис. 4.1, в, г, д). Изменился на этом рубеже не только минеральный состав, но и изотопный: навсегда исчезло независимое от массы фракционирование стабильных изотопов серы (рис. 4.1, б). Появление «озонового щита» в протерозое воспрепятствовало фотолитическому распаду сернистого газа, и изотопная подпись Δ33S практически распрямилась — 0‰. Но озоновый слой мог образоваться только при наличии хотя бы 10−5% кислорода в атмосфере.

Рис. 4.1, б, в, г, д. Независимое от массы фракционирование изотопов серы — показатель наличия озонового слоя (б); содержание осадочного урана в углеродистых сланцах (в); молярное соотношение цинка и железа в карбонатных отложениях (г); молярное соотношение йода и кальция-магния в карбонатных отложениях — показатели насыщенности океана кислородом (д)
К этому же рубежу (2,43–2,24 млрд лет назад) приурочены и первые в истории Земли следы оледенения: тиллиты, дропстоуны и уже упоминавшиеся варвиты. Тиллиты — это глинистая толща, вмещающая хаотически разбросанные угловатые разноразмерные валуны, а дропстоуны — эти самые валуны, часто с ледниковой штриховкой. Образуются подобные отложения на дне морей и озер, куда случайным образом падают обломки горных пород из тающих на поверхности водоема айсбергов и плавучих ледников. Ранее обломки были захвачены на суше при движении по ее поверхности ледового массива, поэтому на крупных валунах остаются приметные параллельные царапины и борозды — следы скольжения мелких обломков.
Если попытаться представить себе мир накануне кислородного события и через несколько миллионов лет после него, скажем на побережье небольшого вулканического острова, то архейская Земля будет выглядеть примерно так: черный пиритовый песок, омываемый волнами темно-сине-зеленого океана, куда впадают желтые серные ручьи, стекающие со склонов невысоких извергающихся вулканов. И весь этот пейзаж освещает красноватый диск солнца, то и дело исчезающий в густой оранжевой дымке. На второй картине песок становится красновато-серым, вулканы подрастают и их вершины покрываются снегом, а ручьи приобретают прозрачность и впадают в мутный бурый океан (в нем окисляется железо), но небо уже голубеет, яснеет, и солнце в зените утрачивает красноту.
Глава 10. Великое кислородное событие, акт второй: атмосфера

Рис. 4.1, н. Палеотемпературы в архейском и протерозойском эонах (3,5–0,5 млрд лет назад)
Великое кислородное событие выразилось не только в накоплении в атмосфере свободного кислорода, но и в изменении состава других газов. Метан с появлением мощного окислителя уже не мог накапливаться в атмосфере. А поскольку одного углекислого газа, который постоянно расходовался на выветривание горных пород, было недостаточно для создания постоянной атмосферной «шубы», температура в приземных слоях атмосферы начала падать (рис. 4.1, н). Более того, в полосчатых железистых формациях и шунгитах захоранивались огромные объемы неокисленного органического углерода, и этот процесс препятствовал возобновлению прежнего уровня углекислого газа, который был изъят из атмосферы при фотосинтезе органического вещества. Если бы не это обстоятельство, Земле, вероятно, была бы уготована участь Венеры, где парниковый эффект, усилившийся по мере разогрева Солнца, привел к испарению океана.
Изменению температурно-газового режима атмосферы на Земле способствовали и два других фактора. Оба они были связаны с ростом площади суши: возросло альбедо (от лат. albus — ‘белый’) планеты («голые» безлесые континенты в отличие от океанов не столько поглощают солнечные лучи, нагреваясь, сколько отражают, остывая), и значительно увеличились площади горных пород, доступные для выветривания — по сути, связывания углекислого газа. Формулу этого процесса, если рассматривать наиболее распространенные горные породы — с высоким содержанием силикатов, — можно представить так:
СаSiО3 + 2СО2 + 3H2O → Са2+ + 2HСО−3 + H4SiО4.
Затем в морской среде H4SiО4, Са2+ и HСО−3 осаждаются как кремнезем и карбонат кальция, с высвобождением СО2, но в меньшем объеме:
H4SiО4 → SiО2 + 2H2O;
Са2+ + 2HСО−3 → СаСО3 + СО2 + H2O.
По счастью, этот процесс — саморегулирующийся: по мере того как суша покрывается снегом и льдом, площадь выходов горных пород, доступных для выветривания, сокращается, а поскольку вулканы и фумаролы продолжают выделять углекислый газ, его доля в атмосфере вновь начинает расти, приходит очередной парниковый цикл, и планета оттаивает.

Рис. 4.1, а. Рост площади континентальной коры в архейском и протерозойском эонах (3,5–0,5 млрд лет назад)
Как раз к началу протерозойского эона континентальные плиты приросли почти на половину своей нынешней площади (рис. 4.1, а), а горы приблизились к современным высотам. Есть ли этому доказательства? Во-первых, 1,8 млрд лет назад в ископаемой летописи появляются эоловые эрги — области подвижных дюн, связанные с деятельностью ветра в сухом климате, что указывает на наличие обширных внутриконтинентальных площадей. Во-вторых, меняется состав вулканических пород, что связано с формированием вулканических полей на суше (такие вулканиты выдает более кислый состав минералов, например, Fe3+/∑Fe > 0,3). В-третьих, иным становится соотношение некоторых изотопов (87Sr/86Sr), редкоземельных металлов (появление позитивной аномалии европия, источником которого тоже является суша) в морских отложениях, а также окислов (возрастает доля К2О по отношению к Na2О, что отражает переход от размыва древних богатых натрием гранитоидов к калиевым гранитам современного типа). Все это свидетельствует о резком усилении стока с суши и росте площадей последней.
Изотопная подпись стронция требует пояснений, поскольку к изотопной истории этого элемента придется обращаться часто. 86Sr поступает из мантии через гидротермы, а более тяжелый радиоактивный изотоп — продукт распада рубидия (87Rb) — выносится с суши в результате разрушения континентальной коры. В архейском океане, пока не сформировались настоящие континенты, изотопную подпись 87Sr/86Sr на отметках 0,701–0,702 оставляла мантия, но 2,2–1,8 млрд лет назад, с усилением притока тяжелого изотопа, «смысл» подписи изменился на более континентальный — 0,704–0,706. (Для сравнения: современное соотношение этих изотопов во всем океане, независимо от солености отдельных морей и заливов, — 0,7092.)
Лик Земли навсегда изменился и обрел современные черты, хотя суша занимала не более 15% площади, а плато приподнялись только до 4000 м. Росли континенты в первую очередь за счет отложений эвапоритов (соли, образующиеся при сильном испарении в мелководных бассейнах) и карбонатов. Например, кратон Каапвааль прирос карбонатным поясом Кэмпбеллранд, 1,5 км мощностью и площадью 500 000 км2, причем среди карбонатов заметную роль играли строматолитовые рифы. И выходит, что в значительной степени континенты обязаны своим происхождением различным существам, без которых не образуются карбонаты и сульфатные разности эвапоритов (с начала фанерозойского эона в построении континентов начали играть важную роль многоклеточные животные в виде раковин, костей и других окаменелостей, а несколько позднее — и высшие растения, древесина и другие, органические ткани которых превращались в каменный уголь).
Прежние мелкие протоконтиненты постепенно нашли друг друга. Примерно 2,7 млрд лет назад кратоны Сан-Франсиску, Каапвааль, Зимбабве, Пилбара и, возможно, Индостанский щит сформировали материк, который иногда называют Зимваальбара. Кола, Карелия и другие части нынешней Северной и Восточной Европы собрались в Балтию, сибирские щиты — в Сибирь (Сибирскую платформу), а разросшийся Канадский щит вместе с некоторыми фрагментами будущей Западной Европы превратился в Лаврентию. Затем Балтия, Сибирь и Лаврентия, предположительно, сложились в Кенорленд (рис. 10.1). Все эти объединения проходили совсем не мирно: при столкновении континентальных масс дыбились горные цепи, а океаны схлопывались и снова открывались. Вероятно, во всех тектонических процессах существовали определенные, не до конца еще вскрытые закономерности. Так, в 1966 г. канадский тектонист Джон Тузо Уилсон на основе распределения горных пород заметил, что Атлантический океан несколько раз открывался и закрывался и его закрытие совпадало с формированием суперконтинента. Эти повторяющиеся явления теперь именуют циклами Уилсона (существование последнего суперматерика — каменноугольно-пермской Пангеи — выявил еще 100 лет назад немецкий гляциолог Альфред Вегенер). Хотя циклы Уилсона не объясняют всего многообразия глобальных тектонических событий, но суперконтиненты, которые собирали воедино более 75% всей континентальной коры, на планете действительно появлялись примерно с интервалом 600–700 млн лет. Скажем, после распада Кенорленда 1,9 млрд лет назад образовалась Нуна. Это эскимосское слово означает землю у северного океана, а выбрано оно потому, что ядро Нуны составляли северные материки — Балтия, Сибирь и Лаврентия. 1,3 млрд лет назад распалась и она, чтобы через 700 млн лет сложилась Родиния — название этого континента происходит от русского слова «родить», буквально «дать начало» другим континентам. Окружал этот суперконтинент суперокеан Мировия. И если причины появления-исчезновения суперконтинентов еще предстоит найти, то само их существование подтверждается целым комплексом независимых доказательств. Так, палеомагнитные данные определяют широтное положение того или иного континента. Тектонические модели обрисовывают взаимную конфигурацию континентальных плит по наличию общих геологических структур и по размещению конвергентных и дивергентных границ. А состав осадочных отложений подсказывает, что, например, цирконы определенного возраста и состава, обнаруженные в Сибири, Северной Америке и Восточной Европе, имеют общий источник и могли единовременно попасть на все материки, только если их пересекал общий горный хребет.

Рис. 10.1. Положение континентов накануне криогеновых оледенений — 1,0 млрд лет назад
Как уже сказано, вся эта материковая масса служила стоком для углекислого газа, что способствовало ослаблению парникового эффекта: метан оказался практически «съеден» кислородом, а Солнце все еще было на 6% холоднее.
И первая ледниковая эра (гляциоэра) — гуронская — не заставила себя ждать. Она наступила 2,43–2,24 млрд лет назад, и с нее, по сути, начинается протерозой. Названа эта эра по ледниковым отложениям полуторакилометровой мощности, сформировавшимся на Канадском щите в районе современного озера Гурон. Слово «эра» подчеркивает, что это не было однократное событие: каждая гляциоэра длительностью в десятки и сотни миллионов лет включала несколько ледниковых периодов, подобных по временным рамкам нынешнему, начавшемуся 2,6 млн лет назад (14 млн лет назад — в Южном полушарии) и еще далеко не закончившемуся, и межледниковий.

Рис. 10.2. Обызвествленные чехлы возможных цианобактерий Girvanella (диаметр 30 мкм); кембрий (510 млн лет); хр. Западный Саян, Хакасия
Затем уровень углекислого газа значительно поднялся. Об этом свидетельствуют сами организмы, жившие в ту пору, 1,4–1,2 млрд лет назад. Так, изотопная подпись углерода из оболочек планктонных эукариот — акритарх — отражает фракционирование изотопов при парциальном давлении двуокиси углерода, в 10–200 раз превышавшем современное. Цианобактерии с обызвествленными чехлами уточняют эту цифру: все-таки не в 200 раз, а скорее в 10 (сегодняшняя атмосфера содержит 0,04% СО2). Именно при содержании этого газа на уровне 0,36% и ниже у цианобактерий возникает механизм концентрации углерода, включающий активный перенос бикарбоната в клетку и его преобразование в двуокись углерода, что сопровождается выделением ионов гидроксила и, как следствие, понижением кислотности среды и осаждением карбоната кальция. Этот минерал и образует известковую оболочку вокруг бактериального чехла (рис. 10.2).
Вновь масштабные оледенения охватили Землю в неопротерозое — в криогеновом (от греч. κρύος — ‘холод’ и γένος — ‘род’, ‘происхождение’) и начале эдиакарского периода. Действительно охватили, судя по распространению ледниковых отложений — тиллитов-дропстоунов-варвитов, а также морозобойных клиньев, следов ледникового выпахивания (параллельных борозд на прочных коренных породах, процарапанных при движении по ним массы льда с захваченными каменными обломками), узорчатых грунтов (полигональной системы трещин, возникших при многократном промерзании и оттаивании грунта) и, конечно, морен (нагромождений валунов и галек, собранных ледником по мере движения). С этими отложениями сопряжены и железные руды, снова полосчатые. Но на этот раз железо поступало с суши, где в межледниковые эпохи выветривались базальты, богатые железом и неодимом. Присутствие последнего и указывает на источник металлов.
Ледниковые шапки простирались от полюсов до экватора: например, в Австралии ледники находились на широте 7,5–8,4 (рис. 10.1, 10.3). Отмечаются три основных оледенения: стертское, маринойское и гаскьеское (720–660, 640–635 и 580 млн лет назад соответственно).

Рис. 10.3. Тиллиты маринойского оледенения; криогений (640–635 млн лет); горы Флиндерс, Южная Австралия
Отложения каждого из трех ледниковых периодов перекрыты маломощным (<5 м) пластом венчающего доломита с резко отрицательным соотношением изотопов углерода. В таком доломите находят полости со сростками-ботриоидами огромных арагонитовых кристаллов (до 20 см длиной), обильные пластовые трещины (полости, теперь, конечно, заполненные морским цементом, между пластами), странные трубчатые структуры, так и названные тубстоунами, а также темпеститы (от англ. tempest — ‘буря’) — отложения, образовавшиеся в результате взламывания ураганами уже затвердевшего карбонатного дна. Если попытаться представить, в каких условиях могли образоваться венчающие доломиты, чтобы в них проявились подобные признаки, можно сделать вывод, что, скорее всего, то были жаркие условия (арагонитовые ботриоиды), когда в океанах зарождались грандиозные торнадо (темпеститы), а на мелководных шельфах быстро таяли метангидраты (кристаллические соединения метана с водой) и по трещинам (тубстоуны и пластовые трещины) в морском дне на поверхность устремлялись потоки метана. Также аномальная изотопная подпись углерода является следствием фракционирования изотопов этого элемента между газом и ионами карбоната при возросшем парциальном давлении СО2 и повысившейся температуре.
Высокий уровень углекислого газа подтверждается и другой изотопной аномалией, характерной для барита (сульфата бария), встречающегося в венчающем доломите, — Δ17О с негативными значениями до −0,70‰, небывалыми для всей последующей истории Земли. Современные значения Δ17О (+5,84‰) в сульфатах обусловлены окислением серосодержащих газов в тропосфере. Для того чтобы этот показатель снизился до −0,70‰, фракционирование должно происходить при аномально высоком парциальном давлении СО2 — в 750 раз выше нынешнего. При таком уровне СО2 тропосфера обогащается стратосферным кислородом с негативным показателем Δ17О, образование молекул которого из озона сопровождается независимым от массы фракционированием изотопов этого элемента. А избыточный углекислый газ как раз мог образоваться за счет окисления метана.
Получается не один парадокс — экваториальные ледники, а все два, включая быструю, в течение нескольких тысяч лет смену ледниковых условий парниковыми. Есть множество гипотез, объясняющих эту совокупность явлений, но нельзя сказать, что причина установлена. Понять механику криогеновых глобальных оледенений, однако, необходимо. Иначе все попытки предсказать даже ближайшее климатическое будущее Земли лишаются смысла.
Гипотезы о причинах криогеновых похолоданий можно разбить на четыре группы: астрофизические, тектонические, физико-химические и биосферные. Первые сводятся в основном к «мечтательным умствованиям» (выражение заимствовано у Венедикта Ерофеева): а что, если у Земли было ледяное кольцо, как у Сатурна? Или: а что, если ось вращения планеты лежала ближе к плоскости ее орбиты (≥54° вместо нынешних 22–24°), как у Урана? В обоих случаях оледенения наступали бы в низких широтах либо из-за тени от кольца, либо от слабого теплового потока вблизи экватора.
Каких-либо способов удостовериться, что «в той степени, в которой научное высказывание говорит о реальности, оно должно быть фальсифицируемо, а в той степени, в которой оно не фальсифицируемо, оно не говорит о реальности» (выражение заимствовано у Карла Поппера), авторами большинства гипотез, увы, не предлагается. Геологические данные, подтверждающие подобные идеи, отсутствуют. Точнее, они есть, но свидетельствуют как раз об обратном. Так, при положении оси вращения Земли под острым углом к плоскости ее орбиты полярная область, обращенная к Солнцу, за день бы прогревалась до точки кипения воды, а в тропиках, пусть и получавших меньше тепла, все равно бы наступала пара «курортных» сезонов — во время весеннего и осеннего равноденствия. Тогда бы ледниковые щиты нарастать не успевали, а центры накопления карбонатов и эвапоритов сдвинулись бы к полюсам. Ничего подобного в ископаемой летописи не наблюдается. Не «выручило» бы и ледяное кольцо: вследствие наклона оси тень от него падала бы лишь на зимнее полушарие, что тоже не способствует глобальному оледенению. Это явление определяется не суровыми зимними морозами, а низкими летними температурами, чтобы зародившийся ледник не успевал растаять. (Не зима должна быть долгой, а лето должно быть коротким.) И не будем забывать об относительной стабильности системы Луна — Земля — Солнце, которая исключает резкие колебания наклона оси планеты, но обеспечивает плавную, с периодом 26 000 лет, смену положения оси вращения планеты — прецессию. Прецессия тоже смягчала бы зимы при наличии ледяного кольца.
Тектонические гипотезы предполагают такие взаимные конфигурации континентов, которые хотя бы отчасти объясняли развитие на них ледниковых отложений. Увы, самое простое из возможных объяснений — сосредоточие континентальных масс вблизи полюсов, подобно современной Антарктиде или древней Пангее, — не подтверждается: суперконтинент Родиния располагался так, что значительные области суши оказались вблизи палеоэкватора. К концу криогенового периода этот суперконтинент как раз начинал дробиться (отделилась Лаврентия). Поэтому есть возможность увязать образование приэкваториальных ледников с зонами дробления — рифтами, подобными Восточно-Африканской рифтовой системе, вдоль которой растут вулканические цепи, и вершины самых высоких гор (Килиманджаро) покрыты снежниками. Однако площадные оледенения, в том числе следы обширных шельфовых ледовых полей (в виде отложений тиллитов, достигающих нескольких сотен метров мощности), в линейные рифтовые структуры никак не вписываются, и гипотеза «Земли на рифтовых молниях» тоже отпадает. Эта модель больше подходит для гуронской гляциоэры и отчасти для маринойского оледенения.
Климатические модели типа «Земля — талый комок» связывают площади горных пород, подвергающихся выветриванию, их состав и влияние наземной биоты на скорость выветривания с параметрами гидросферы (стратифицированный или смешанный, но обязательно открытый в области экватора океан, а в последнем случае и наличие морских течений определенной направленности) и атмосферы (газовый состав). Эти модели достаточно хорошо объясняют, что ледяной щит средней мощности 2500 м (и до 5000 м в Лаврентии и Антарктиде — этот континент часто оказывался крайним) мог вырасти за несколько сотен тысяч лет. Ледники зародились в Гренвильской горной системе (ее остатки составляют восточную часть Канадского щита), достигавшей высоты 2000 м, и за 50 000 лет охватили весь суперконтинент Родинию; уровень Мировии упал на 200 м. Однако венчающий доломит с этой моделью согласуется плохо.
Модель «Земля — снежок», предложенная американским геофизиком Джозефом Киршвинком в 1992 г., вообще предполагает длительное суровое оледенение всей планеты при резких скачках уровня углекислого газа и относительно неплохо увязывает чередование ледников в тропических широтах с быстрым образованием на их месте венчающего доломита. Важная роль в этой модели отводится суперконтиненту, низкоширотное положение которого повышает альбедо планеты, а обширная суша служит стоком углекислого газа — так начинается холодный период. Стоит же ледникам спуститься от полюсов до широт 30°, образование «снежка» неизбежно, поскольку альбедо снега и льда еще выше, чем у голого континента. Когда же вся поверхность суши покрывается льдом, углекислый газ перестает расходоваться на выветривание и начинает накапливаться в атмосфере (вулканическая деятельность ведь не прекращается). И достаточно парциальному давлению СО2 достичь уровня 12 000 Па (для этого при современных темпах поступления двуокиси углерода требуется всего несколько миллионов лет), весь процесс разворачивается в обратную сторону и развивается по нарастающей вплоть до образования венчающего доломита, знаменующего приход очередной парниковой эры. Как показали симуляционные модели, весь ледовый панцирь, учитывая низкоширотное положение ледников, может растаять всего за несколько тысяч лет.
Конечно, сплошные оледенения не длились десятки миллионов лет, а прерывались потеплениями, о чем свидетельствует чередование осадочных отложений разного происхождения. В стертских отложениях Австралии, например, распознается четыре ледниковья, перемежавшихся с теплыми эпохами. Но как жизнь сохранялась под сплошным ледяным панцирем? Вероятно, в гигантских трещинах во льдах, которые раскалывались в районах выхода гидротермальных источников или при торошении, вызванном лунными приливами. Одни организмы существовали в подледных озерах, подобных современному озеру Восток в Антарктиде. Другие выживали в пузырьках воздуха во льду, как некоторые одноклеточные на Байкале, пережидающие суровые зимы от оттепели до оттепели. Третьи — в криоконитовых ямах, которые протаивали под скоплениями темной минеральной пыли — криоконита (такие ямы известны на всех крупных ледниках). Неудивительно, что на кратоне Сан-Франсиску в ленточных глинах обнаружено от 1,7 до 4% органического вещества. Может быть, такое «скучивание» организмов на небольших площадях даже сыграло положительную роль в эволюции — именно в таких оазисах могли зародиться многоклеточные.
Остается необъяснимым, или необъясненным, лишь один пунктик: что провоцировало начало ледниковых эр? Если исключительно положение континентов, то почему между гуронской и криогеновой гляциоэрами (более миллиарда лет) глобальные оледенения не случались?
Впрочем, если на Земле в то время бурно эволюционировали эукариоты, то их эволюция должна была привести к появлению массы фитопланктона, который, в отличие от цианобактерий, производит больше органического вещества на единицу потребленного азота и фосфора. А чтобы расти, все эти одноклеточные водоросли должны были фотосинтезировать, захватывая углекислый газ. Превращаясь со временем в мортмассу, огромные объемы органического вещества уходили в бескислородные толщи океана, начинавшиеся уже на небольшой глубине, унося углерод и не давая ему окисляться и вновь возвращаться в атмосферу в виде двуокиси. В связи с этим содержание парникового газа в атмосфере могло снизиться до критического, что и привело к наступлению холодов.
Так ли было на самом деле? Эволюция фитопланктона, главным образом акритарх, неплохо запечатлена в геологической летописи: их разнообразие с начала протерозойского эона ко времени тропических оледенений возрастало почти по экспоненте (рис. 4.1, м). Однако для поглощения значительных объемов углекислого газа важно не разнообразие, а общая биомасса и размер отдельных особей (крупные формы погружаются быстрее, особенно когда слипаются вместе). Для набора биомассы требуется только подкормка (равно как и для набора массы). Косвенными свидетельствами усилившегося в это время потока органического вещества на дно служат неопротерозойские сдвиги изотопной подписи углерода в карбонатных отложениях в сторону резко негативных значений (до −10‰), предшествовавшие каждому оледенению. Поскольку в бескислородной обстановке органика разлагалась анаэробными бактериями, фракционирование ими изотопов углерода и могло выразиться в этих негативных аномалиях. В закисном океане органическое вещество разлагается благодаря восстановлению железа и сульфата, в результате чего образуется не столько двуокись углерода, сколько бикарбонат-ион, который, реагируя с железом, выпадает в осадок в виде сидерита:
2CH2O + 8Fe3+(OH)3 → 8Fe2+ + 2HCO3− + 14OH− + 6H2O;
8Fe2+ + 8CO32−→ 8FeCO3.
Получившийся сидерит и формировал полосчатые железистые руды криогенового периода.
Для подкормки фитопланктона дефицитным фосфатом свою лепту могла внести и древняя наземная биота, которая, судя по некоторым седиментологическим признакам, уже существовала. Одним из показателей разрушительно-созидательной деятельности наземной биоты, т. е. химического выветривания, как раз и является рост содержания фосфора в морских отложениях. С криогенового периода (800 млн лет) массовая доля фосфора в ископаемой летописи (конкретно, в сланцах) возрастает сразу в 4 раза — с 0,051 до 0,209%. Увеличивается и число фосфоритовых месторождений. Вероятно, наземную биоту представляли некоторые группы актинобактерий, водорослей, грибов и лишайников, которые химически и механически взламывают кристаллическую решетку. Лишайники являются симбиозом все тех же водорослей и грибов и, несмотря на свой невзрачный вид — корки на камнях, небольшие рожки и «кустики», — заметно повышают темпы химического выветривания. Актинобактерии — древняя группа прокариот — способны извлекать ионы Mg, Ca и Si из базальтов, коматиитов и гранитов в два-три и даже в десятки раз быстрее (в зависимости от породы), чем любые химические процессы.
Пока следы достаточно сложной наземной биоты, существовавшей 1,0 млрд лет назад, обнаружены только в Северо-Западной Шотландии: в озерных отложениях того времени, которые ныне слагают песчаные скалы Торридона, есть прослои серых сланцев, а в них — фосфатные стяжения с микроскопическими остатками. Несмотря на мелкие размеры, некоторые из ископаемых намного крупнее любых бактерий. Среди них встречаются и многоклеточные формы, и клетки с очень сложным строением оболочки (рис. 10.4). Вероятно, то были одноклеточные и колониальные водоросли, а росли они вместе с колониальными коккоидными и нитчатыми цианобактериями на дне временно пересыхающих водоемов.

Рис. 10.4. Ископаемые организмы, возможно цианобактерии и эукариоты, в озерных отложениях; мезопротерозой (1,2–1,0 млрд лет); Торридон, Шотландия (публикуется с разрешения Мартина Брэзие); а — парные и строенные клетки (диаметр 10 мкм); б — созревшая циста (диаметр 30 мкм)
В океане на биогенах бурно развивался фотосинтезирующий планктон, и образовавшееся органическое вещество со временем уходило на дно. О высоких темпах захоронения неокисленной органики свидетельствует углеродная изотопная аномалия, приуроченная к морским отложениям, как раз предшествующим началу стертского оледенения (720 млн лет назад): δ13С = +13‰. И если органика погребается в неокисленном виде, то весь углекислый газ, выхваченный из атмосферы и связанный в виде органических соединений, не поднимется вновь в воздух, а его содержание может упасть ниже критической отметки, при которой поддерживается парниковый эффект. Наступает похолодание. Этот процесс саморегулирующийся: похолодание вызывает вымирание планктона, двуокись углерода больше не изымается из атмосферы (и не расходуется на выветривание скованных ледяным панцирем горных пород) и начинает накапливаться в атмосфере, поступая вместе с вулканическими эманациями. Парниковый эффект возобновляется, льды тают, талые воды выносят в океан огромные объемы накопившихся во льдах в виде пыли биогенов, фитопланктон «цветет»... Судя по многочисленности оледенений в криогеновом-эдиакарском периодах (по меньшей мере три, а возможно и пять, не считая более мелких циклов), биосферная гипотеза наиболее близка к истине.
Глава 11. Великое кислородное событие, акт третий: биосфера
Наконец, третье и наиболее существенное последствие Великого кислородного события — появление эукариот, что, собственно, привело ко второму событию (изменению состава атмосферы) и предопределило первое (распространение новых минералов). Вполне возможно, что случилось это событие вынужденно: свободный кислород — вещество исключительно активное и легко взаимодействует с органическими молекулами, окисляя их. Поэтому избыток кислорода грозит живым существам гибелью. Выход из ситуации был найден неожиданный и радикальный: создание своего рода живой матрешки, где внутри одной клетки, изолированно от ее внутренних структур, с помощью дополнительной мембраны была упрятана другая. Так предшественники эукариот обрели симбиотическую органеллу митохондрию, обеспечивающую защиту от опасного кислорода благодаря его постепенному усвоению, т. е. дыхание. У митохондрий до сих пор сохранилась собственная ДНК, указывающая на их прямое родство с альфа-протеобактериями, которые используют кислород как акцептор электронов для пополнения энергетических запасов клетки, а именно для синтеза аденозинтрифосфорной кислоты (АТФ). Симбиогенез с альфа-протеобактериями обеспечил будущих эукариот необходимыми запасами дешевой энергии. Ведь из всех возможных путей обмена веществ именно кислородное дыхание дает наибольший выход свободной энергии в расчете на одну использованную (окисленную) молекулу органического вещества (например, глюкозы). Без подобной энергетической подпитки никогда бы не появились ни подвижные существа, ни многоклеточные, ни крупные одноклеточные вообще. Даже организмы, перешедшие на анаэробное дыхание, по-прежнему пользуются митохондриями, хотя у них эти органеллы преобразовались в гидрогеносомы и митосомы, восстанавливающие электроны до водорода. (Не исключено, что в этом случае бывшие альфа-протеобактерии «вспоминают» свое прошлое: так, пурпурные несерные бактерии, относящиеся к этой группе, используя разные источники протонов, могут жить и в аэробной, и в анаэробной среде.) За длительное, по меньшей мере 2 млрд лет, время сосуществования с хозяевами митохондрии передали значительную часть своего генного аппарата клеточному ядру, и в ядерном геноме оказались чужеродные гены все тех же альфа-протеобактерий, например такие, что кодируют белки, устойчивые при высоких температурах.
Вслед за этим актом обретения сложности последовал еще один: клетки с митохондриями обзавелись собственными фабриками фотосинтеза — пластидами, или хлоропластами. Последние являются прямыми потомками свободно живущих цианобактерий. Среди возможных претендентов на эту роль рассматриваются и шаровидные хроококки. Хроококки улавливают в дневное время азот, для чего используют запасенные ночью полисахариды и крахмал (многие цианобактерии на это не способны). Обретение фотосимбионта оказалось выгодным вдвойне: сразу и органические запасы, и азотистые «удобрения». Возможно, именно потребность в фиксации азота — значимого биогенного элемента — могла быть первичной задачей симбиогенеза. Организм-хозяин усваивает его в виде аммонийного иона (NH4+). Так появились красные и зеленые водоросли — первичнопластидные, или архепластиды (Archaeplastida).
Как и в случае с митохондрией, часть генетической информации новой органеллы была передана ядру, геном которого у растений содержит почти 20% генов цианобактерии (а пластиды удержали менее 10% изначального бактериального генома).
Хотя теория симбиогенеза — многоступенчатого симбиоза различных прокариот, сформировавших эукариотную клетку, — зародилась более 100 лет назад — в трудах российских ботаников и физиологов Андрея Сергеевича Фаминцына, Константина Сергеевича Мережковского и Бориса Михайловича Козо-Полянского, их правоту подтвердили только молекулярно-генетические исследования последних лет. Более того, если до нынешнего тысячелетия считалось, что строение клеточных мембран не позволяет одному микробу поглотить другого, чтобы тот остался цел и невредим, недавнее открытие протеобактерий двух разных групп, которые существуют одна внутри другой, делятся продуктами обмена веществ и даже обмениваются генами, доказывает возможность симбиоза разных прокариот.
В железорудной формации Ганфлинт (1,9 млрд лет) наряду с простенькими бактериями найдена довольно сложная эосфера (Eosphaera), представляющая собой относительно крупную клетку (до 30 мкм) с двойной оболочкой, между слоями которой закономерно расположены мелкие сферические тельца (1–7 мкм в диаметре). Даже в более древней формации Стрелли-Пул (3,43 млрд лет) присутствуют относительно большие и сложные по меркам микромира чечевицеподобные клетки — с оторочкой на поверхности и мелкими сферическими тельцами внутри. Так вполне могли выглядеть древние симбиотические прокариоты, пока еще не превратившиеся в эукариот. А в мелководных морских аргиллитах и сланцах возрастом 3,2 млрд лет на Каапваале найдены органические сферы до 0,3 мм в диаметре — самые крупные организмы архейского эона. Они намного прочнее бактериальных оболочек, но в то же время не имеют сложной структуры стенки и намного крупнее большинства бактерий. И такими вполне могли быть начинающие эукариоты.
Что касается клетки-хозяина — той, которая гостеприимно предоставила кров альфа-протео- и цианобактериям, — то ею могла послужить асгардархея. Эта группа архей, представители которой носят имена древнескандинавских богов (а вся группа — имя их святилища), наиболее близка к эукариотам по молекулярным данным и обладает множеством генов, отсутствующих у всех прочих прокариот, но характерных для эукариот. Например, у них есть гены, кодирующие белки для построения цитоскелета, передачи химических сигналов, образования и перемещения вакуолей внутри клетки. По характеру обмена веществ и строению клеточной мембраны асгардархеи тоже близки к эукариотам. (Впрочем, мнения о конкретных участниках и порядке событий симбиогенеза очень разнятся.)
Началу Великого кислородного события предшествовало еще одно важное явление — формирование в палеопротерозое (2,415 млрд лет назад) в Каапваале богатейшего марганцевого месторождения, где содержание Mn достигает 4 × 103 млрд кг. (Марганцевые руды этого времени разведаны также на Индостанском и Западно-Африканском щитах.) Марганец здесь связан с карбонатом — кутнoгоритом [(CaMn2+0.7Mg0.3)CO3)2]. Кутногорит образовался в результате диагенетического восстановления окисла марганца — браунита [Mn3+6Mn2+(SiO4)O8]. Все же литологические и изотопные особенности марганцевой руды указывают на то, что сами первичные окислы кристаллизовались в бескислородной среде! Сильные окислители — свободный кислород, перекись водорода или иные перекиси — тогда просто не существовали. Значит, роль окислителя сыграло что-то другое. Это «что-то» существует до сих пор в виде водоокисляющего комплекса фотосистемы II. В данном комплексе четыре атома Mn, связанных через кислородный мостик с одним атомом Са, являются переносчиками электронов и под действием света способны окислить две молекулы воды до кислорода. Получается, что важнейший комплекс фотосистемы II, который, собственно, обеспечивает пополнение атмосферы кислородом, сформировался еще у анаэробных предшественников цианобактерий, которые и положили начало кислородной революции. Учитывая насыщенность архейского и палеопротерозойского океана ионами Mn2+, которые относительно легко расстаются с электронами, использовать такой ресурс было можно и нужно.

Рис. 4.1, л. Морфологическое разнообразие строматолитов
С появлением свободного кислорода возникли новые группы прокариот, полагавшиеся на кислород как на главный источник энергии, и анаэробным старожилам пришлось потесниться. В конечном счете возросло разнообразие сообществ, что, например, выражается в появлении 2,25–2,05 млрд лет назад множества новых форм строматолитов (рис. 4.1, л). Время событий симбиогенеза можно определить благодаря ископаемой летописи. Альфа-протеобактерия стала симбионтом не позднее 1,8 млрд лет назад: к этому уровню приурочены древнейшие остатки эукариот. С этого же — палеопротерозойского — времени начинают встречаться ископаемые остатки, в которых узнаются сложные организмы — эукариоты. 1,8–1,65 млрд лет назад возникли одноклеточные зеленые водоросли, известные как акритархи. 1,56 млрд лет назад уже существовали крупные (до 30 см длиной и 8 см шириной) многоклеточные водоросли с хорошо выраженным листовидным талломом (слоевищем), 1,2 млрд лет назад — красные водоросли, мало отличимые от современных колониальных бангиевых: длинные цепочки из радиально и поперечно делящихся клеток, окруженных общей внешней оболочкой. 850–650 млн лет назад настало время многоклеточных зеленых водорослей с сифонокладальным талломом (многоядерные клетки, соединенные в ветвящиеся нити) и сифоновых желто-зеленых (кустистая на вид, но одна многоядерная клетка). Все это очень разные группы: зеленые и красные водоросли, как уже сказано, принадлежат к архепластидам, желто-зеленые относятся к страменопилам (Stramenopiles) — другой главной ветви эукариот, представители которой имеют жгутик сложного строения и митохондрии с трубчатыми выступами стенки — кристами; форма крист и подсказала их общее название (от лат. stramentum — ‘соломка’, или ‘трубочка’, и pilus — ‘волос’).
В основном такие органические остатки представляют собой устойчивые к разложению клеточные оболочки, усиленные пигментом. (Пигмент был необходим древним планктонным и мелководным водорослям, чтобы обезопасить себя от ближнего ультрафиолетового и видимого света; более коротковолновое излучение уже экранировалось озоном и кислородом.) Многие из этих находок далеко не единичны: например, сатка (Satka), получившая имя уральского городка, тимофеевия (Timofeevia), названная в честь известного исследователя древнейших организмов геолога Бориса Васильевича Тимофеева, и якутиянема (Jacutianema), впервые обнаруженная в Якутии, теперь известны на Шпицбергене, в Австралии, Канаде, Индии и Китае — везде, где имеются сходные морские отложения. Видимо, удачные эволюционные новшества распространялись очень быстро.
Если некоторые из перечисленных групп имеют достаточно выразительную морфологию, позволяющую определить их принадлежность, с акритархами все было очень непросто. Открытые в 1830-е гг. в нижнепалеозойских отложениях, они долгое время считались организмами неясной природы (отсюда и название: от греч. α-κρϊτος — ‘неопределенный’ и άρχή — ‘начало’). Причем известно их было не так уж много. Лишь столетие спустя, когда Б. В. Тимофеев нашел способ извлекать многочисленные органические оболочки акритарх из аргиллитов и алевролитов с помощью смеси сильных кислот, эти ископаемые превратились в наиболее важные индикаторы возраста докембрийских отложений и обстановки, в которой они формировались. Было понятно, что эти миниатюрные сморщенные диски (не более 0,8 мм в диаметре) когда-то были шариками, плавающими в водной толще, — фитопланктоном. Но только новейшие методы микроскопии и анализ состава вещества позволили отчасти раскрыть загадку их принадлежности. Шарики оказались упакованы в очень устойчивую (к воздействию кислот) трехслойную клеточную оболочку, в составе которой выявлены полиароматические и разветвленные алифатические углеводороды, адсорбционные спектры которых ближе всего к альгенанам зеленых водорослей. Похоже, что у акритарх чередовались половое и бесполое поколения. От представителей последнего сохранились гладкие, легко распадавшиеся оболочки цист, покоившихся на дне. Каждая циста, вероятно, содержала зиготу, образовавшуюся при слиянии половых клеток. У цист хорошо выражен пилом — щель разверзания, через которую зооспоры или автоспоры — результат деления зиготы — когда-то покинули цисту. Размножавшиеся половым путем (на что опять же указывает наличие пилома), но нередко и делением, особи имели сложные оболочки с многочисленными оторочками, шипами или выростами, видимо удерживавшими слизь. Все эти поверхностные структуры повышали плавучесть. Внутри такой оболочки и находилась циста.
Именно акритархи преобладали среди протерозойских планктонных эукариот как по числу видов, так и по количеству экземпляров, и впоследствии изобиловали в палеозойских морях наряду с празинофитовыми (такими, как Tasmanites). Празинофитовые отличаются от акритарх по характерным пористым органическим оболочкам. Менее распространенными планктонными зелеными водорослями были одноклеточные существа с ажурными, сетчатыми, округлыми фосфатными скелетиками (Characodictyon и другие, 15–50 мкм в диаметре). Ныне подобные фосфатные чешуйки тоже есть только у празинофитовых. Пока такие скелетики известны лишь из относительно глубоководных отложений (группа Фифтинмайл), сформировавшихся 800–700 млн лет назад на границе нынешней Аляски и Канады.
Особенно бурный рост водорослевого фитопланктона начался после стертского оледенения. Это событие фиксируется по значительному сдвигу в соотношении биомаркеров: до оледенения ископаемое органическое вещество было в основном сложено гопанами, а после более половины объема такого вещества стали составлять стераны. Первые являются продуктами распада бактериальных остатков, а вторые — эукариотных. Интересно, в свою очередь, соотношение разных молекул среди стеранов: холестаны, преобладающие в достертских отложениях, свидетельствуют о том, что в океане того времени обитали красные водоросли и гетеротрофы, а послестертские эргостаны и стигмастаны могли принадлежать зеленым водорослям. Следовательно, с послестертского времени (около 650 млн лет назад) эукариоты становятся важной группой в океане. Сам же стремительный рост разнообразия и биомассы фитопланктона был обусловлен тем, что таяние ледников усилило приток в океан биогенных веществ.
Одновременно менялось и донное сообщество. Бактериальные пленки и даже биоматы были двумерными, поскольку даже вертикальные нити не добавляли бактериальным сообществам прижизненной пространственной структуры (строматолиты — это все-таки посмертные монументы). Бентосные зеленые, желто-зеленые и красные водоросли благодаря своим размерам образовали на дне морей первые трехмерные сообщества организмов — подводные луга. Такие луга распространились в конце мезопротерозойской — начале неопротерозойской эры (1200–650 млн лет назад), несмотря на суровое криогеновое похолодание (850–650 млн лет назад): ископаемые остатки «луговой растительности» известны из Сибири, Канады, Шпицбергена и Южного Китая. Количество видов в отдельных сообществах достигало 30 и более (в палеопротерозое не доходило и до 10). Однако стабилизаторами подвижного субстрата, склеивавшими частицы осадка в плотный грунт, пригодный для закрепления водорослей, все еще выступали нитчатые цианобактерии, чьи остатки достаточно обычны среди позднепротерозойских комплексов.
Обилие отмершей органики не могло не привести к появлению крупных ее потребителей. Бактерии, такие как сульфатвосстановители (чье присутствие выдают микроскопические цепочки пирита), хотя и справляются с компостом, но не так споро. Им на помощь к концу мезопротерозойской эры пришли одноклеточные грибы. Одни из них были похожи на хитридиевые (Chytridiomycota), другие — на мукоровые (Mucoromycota). Протерозойские хитридиевые хорошо определяются по характерным зооспорангиям — овальная клетка с оболочкой и ризомицелием в виде тонких ветвящихся нитей, которые врастают в субстрат — в отмершие талломы водорослей. Спорангиеносцы древних мукоровых, как и современные, похожи на миниатюрные спички. Правда, у «спички» бывает сдвоенная «головка» — сидящие друг над другом спорангии: в нижнем формировались многочисленные споры (он практически непрозрачный), верхний содержал всего несколько спор (он прозрачный с различимыми отдельными клетками). Вероятно, разнообразие протерозойских грибов было очень большим. Например, среди акритарх есть оболочки, покрытые трубочками с поперечными перегородками, очень похожими на септированные грибные гифы — тонкие трубочки, образующие грибницу. Встречаются сложные сплетения гифов, видимо принадлежавших многоклеточным грибам. А самые древние возможные грибы, возрастом 2,4 млн лет, происходят из Каапвааля: в полостях, образовавшихся в подушечных базальтовых лавах, они росли в виде мицелия из тонких (2–12 мкм в диаметре) полых переплетающихся нитей.
Учитывая, что грибы имели общих с животными прародителей, время их появления косвенно определяет временной рубеж, не ранее которого должны были возникнуть и существа, давшие начало многоклеточным животным, — около 1 млрд лет. Вместе грибы, многоклеточные животные и некоторые одноклеточные организмы составляют единую группу заднежгутиковых эукариот (Opisthokonta), поскольку хотя бы на одной стадии жизненного цикла у них появляются клетки с одним двигательным жгутиком. Грибы, эволюционируя как сидячие осмотрофы, потеряли часть «ненужных» им предковых генов, сохранившихся у животных.
Пока же из криогеновых отложений известны только одноклеточные представители других гетеротрофов, иначе говоря, хищников. Это раковинные, или лобозные, амебы (Amoebozoa), фораминиферы с тонкими ложноножками — ризоподиями (Rhizaria) и сидячие инфузории-тинтинниды (Alveolates, от лат. alveolus — ‘желоб’). Для клеток альвеолят характерным признаком является слой пузырьков (альвеол), расположенный непосредственно под клеточной мембраной. Это значит, что не позднее криогенового периода на Земле уже существовали все основные группы эукариот: архепластиды, страменопилы, альвеоляты, ризарии, амебозои и опистоконты. (Не хватает лишь экскават, но эти крошечные одноклеточные паразитические существа практически не имеют шансов попасть в палеонтологическую летопись.) Различаются остатки древних одноклеточных раковинных организмов по составу, форме и микроструктуре раковинок и органического вещества. У древних фораминифер (Foraminifera), например, раковинка представляет собой пористую трубочку с агглютинированной стенкой, т. е. состоит из мельчайших зерен кварца, слюды, гематита и полевого шпата (эти же минералы слагают вмещающую породу, но в породе зерна разноразмерные и среди них много крупных частиц); поры были необходимы для выхода ризоподий, которыми фораминифера захватывала добычу. Питались все эти существа другими, более мелкими одноклеточными, от которых совсем ничего не осталось. Или пока ничего не нашли.
Выход на сцену одноклеточных хищников не мог не преобразить всю биоту. (В современных водоемах внедрение выедателей резко усиливает темпы роста зеленых водорослей и диатомовых, поскольку выедание планктона ускоряет круговорот ранее потребленных питательных веществ. В новых поколениях водорослей увеличивается содержание белков, что делает их еще более привлекательными для гетеротрофов. Наоборот, количество цианобактерий уменьшается, хотя они и защищаются от выедателей с помощью ядов.)
И тогда, между 800 и 540 млн лет назад, роль строматолитов резко уменьшилась (рис. 4.1, л). Значительно сократилось не только разнообразие построек, но и обилие: все реже формировались отложения при участии строматолитовых сообществ. Поскольку коренная перестройка в химическом составе океана еще не произошла, конец строматолитовой эпохи мог наступить из-за развития фитопланктона, препятствовавшего проникновению света в придонные слои. Отчасти этому способствовали и появление бентосных эукариотных «лугов», развивавшихся быстрее цианобактериальных сообществ и занимавших их место, и системообразующая роль выедателей.









