Позднепермского эунотозавра лишили звания «предка черепах»

Происхождение черепах — биологическая загадка, волнующая ученых на протяжении долгих десятилетий. Молекулярные биологи уже давно выдвинули предположение, что ближайшие родственники черепах — архозавры (крокодилы, динозавры и птицы). Однако палеонтологические исследования эту гипотезу не подтверждали. Использовав метод рентгеновской компьютерной микротомографии, международная команда исследователей изучила остатки нескольких предполагаемых предков черепах, в том числе эунотозавра, жившего в Южной Африке около 260 миллионов лет назад. Эунотозавр оказался морфологически гораздо дальше от черепах, чем было принято считать до сих пор — судя по всему, его «черепашьи» черты развились конвергентно. А вот версия родственных связей черепах и архозавров получила морфологическое подтверждение: в черепе проганохелиса, несомненной ранней черепахи, удалось обнаружить черты сходства с архозаврами.
Черепахи — одни из самых необычно выглядящих современных рептилий. Во-первых, у них анапсидные (то есть лишенные височных окон) черепа, что резко отличает их от диапсидных черепов других современных рептилий, у которых таких окон два. Во-вторых, их посткраниальные скелеты превратились в крепкие панцири: ребра, позвонки и кожные окостенения слились в два щита брони, верхний (карапакс) и нижний (пластрон), причем большая часть карапакса произошла именно от расширенных скелетных ребер (Hirasawa et al., 2013. The endoskeletal origin of the turtle carapace). Наконец, заключение самих себя в подобную броню, неотделимую от туловища, вынудило черепах перестроить плечевой пояс, так что, в отличие от других позвоночных, их лопатки находятся внутри грудной клетки, а не снаружи (Nagashima et al., 2012. Body plan of turtles: an anatomical, developmental and evolutionary perspective), и «изобрести» уникальный способ дыхания, при котором для расширения легких используются специальные мышцы, связанные с внутренними органами и конечностями (Lyson et al., 2014. Origin of the unique ventilatory apparatus of turtles).
В общем, не стоит удивляться, что вопрос об их происхождении оставался одним из самых запутанных в эволюционной биологии. В разное время этих панцирных пресмыкающихся то относили к «примитивным» рептилиям-анапсидам, то называли их ближайшей родней ящериц и змей, то помещали в качестве сестринской группы по отношению к архозаврам (O. Rieppel, M. deBraga, 1996. Turtles as diapsid reptiles; T. R. Lyson et al., 2011. MicroRNAs support a turtle + lizard clade; N. Iwabe et al., 2004. Sister Group Relationship of Turtles to the Bird-Crocodilian Clade Revealed by Nuclear DNA–Coded Proteins). Не было единства мнений и среди палеонтологов, которые искали «черепашьи» черты у самых разных животных и в причисляли к предкам черепах то примитивных проколофонов, тоже обладателей анапсидного черепа, то морских завроптеригий, среди которых были черепахоподобные панцирные плакодонты (R. R. Reisz, M. Laurin, 1991. Owenetta and the origin of turtles; M. deBraga, O. Rieppel, 2008. Reptile phylogeny and the interrelationships of turtles).
Одной из наиболее признанных «стволовых черепах» долгое время считался эунотозавр (Eunotosaurus africanus), обнаруженный в южноафриканской пустыне Кару в конце XIX столетия (T. R. Lyson et al., 2010. Transitional fossils and the origin of turtles). Его расширенные ребра и форма позвонков напомнили исследователям ранних черепах. Однако были и причины сомневаться в близком родстве эунотозавра с черепахами. Другие его анатомические черты — например, зубы на костях основания черепа — были не похожи вообще ни на каких других рептилий. Вдобавок, как выяснилось позднее, «черепашьи» черты эунотозавра оказались в той же степени характерны и для ранних рептилий миллереттид (Millerettidae), так что поневоле вставал вопрос, действительно ли он родич черепахам, или обрел сходство с ними независимо (С. E. Gow, 1997. A reassessment of Eunotosaurus africanus Seeley (Amniota: Parareptilia)). Если молекулярные биологи подчеркивали сходство черепах с высшими рептилиями — ящерицами и архозаврами, — то при попытке построить родословную черепах от эунотозавра их «выбрасывало» за пределы эволюционного древа современных рептилий (G. S. Bever et al., 2015. Evolutionary origin of the turtle skull).
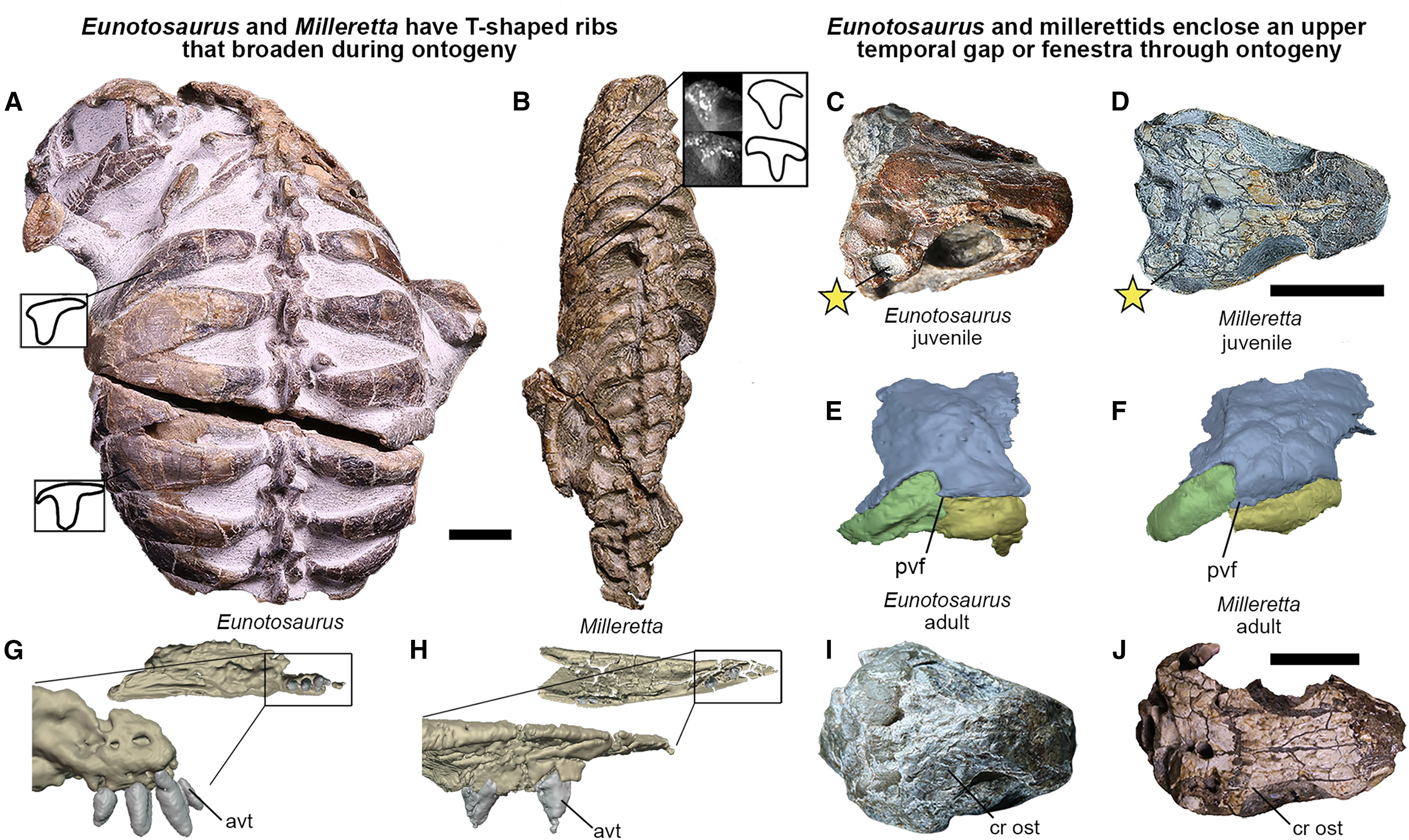
Т-образные в сечении, расширенные рёбра, ранее считавшиеся чертой, сближавшей эунотозавра с черепахами (у которых карапакс, напоминаю, образован в основном расширенными ребрами), характерны и для миллеретты (Milleretta rubidgei) — примитивной рептилии из позднепермских отложений ЮАР. Обратите внимание на сходство черепов эунотозавра (С, E, I) и миллеретты (D, F, J), сохраняющееся как у молодых, так и у взрослых особей. Длина масштабных отрезков — 1 см. Рисунок из обсуждаемой статьи
И вот группа исследователей из США, ЮАР, Франции, Великобритании и Германии решила провести комплексный анатомический и филогенетический анализ и оценить родство эунотозавра с другими предполагаемыми стволовыми черепахами. Ученые собрали самую большую на сегодня морфологическую матрицу, проанализировав для 226 видов ископаемых черепах и их предполагаемых родственников 962 признака. Результаты однозначно продемонстрировали, что строение эунотозавра гораздо сильнее сближает его с примитивными рептилиями миллереттидами, чем с более продвинутыми рептилиями — включая и несомненных ранних черепах вроде позднетриасового проганохелиса (Proganochelys), жившего около 210 миллионов лет назад.
К примеру, если у проганохелиса, как и у остальных высших рептилий, в передней части слуховой капсулы черепа имеются передне-нижний отросток (anterior inferior process) и проотический гребень (crista prootica), а боковые стенки мозговой коробки формируют парные кости — латеросфеноиды, то у эунотозавра и миллереттид ничего подобного нет. Что же касается «черепашьих» черт эунотозавра, то они, вероятно, возникли в результате конвергентной эволюции: южноафриканский ящер, скорее всего, был роющим животным, так что расширенные ребра уберегали его внутренние органы от чрезмерного сдавливания.
Таким образом, черепа ранних несомненных черепах оказались ближе к черепам родственников архозавров, таких как триасовая пролацерта (Prolacerta broomi), чем к черепу эунотозавра, их предполагаемого «предка». Судя по результатам филогенетического анализа, предки черепах отделились от предков архозавров еще в позднем пермском периоде, около 255 миллионов лет назад. У них как минимум 25 общих уникальных черт строения, в том числе небольшой латеросфеноид, обрамляющий эпиоптическое окно (fenestra epioptica; отверстие в глазнично-височной области черепа), и шейные позвонки в форме параллелограмма. Еще в 2015 году на основе молекулярных исследований была предложена клада Archelosauria, объединяющая обе группы (Crawford et al., 2015. A phylogenomic analysis of turtles).
Альтернативные гипотезы о происхождении черепах оказались менее вероятными (требовали большего количества «шагов» на эволюционном древе) по сравнению с «архозавровой» версией, поэтому авторы их отвергли.
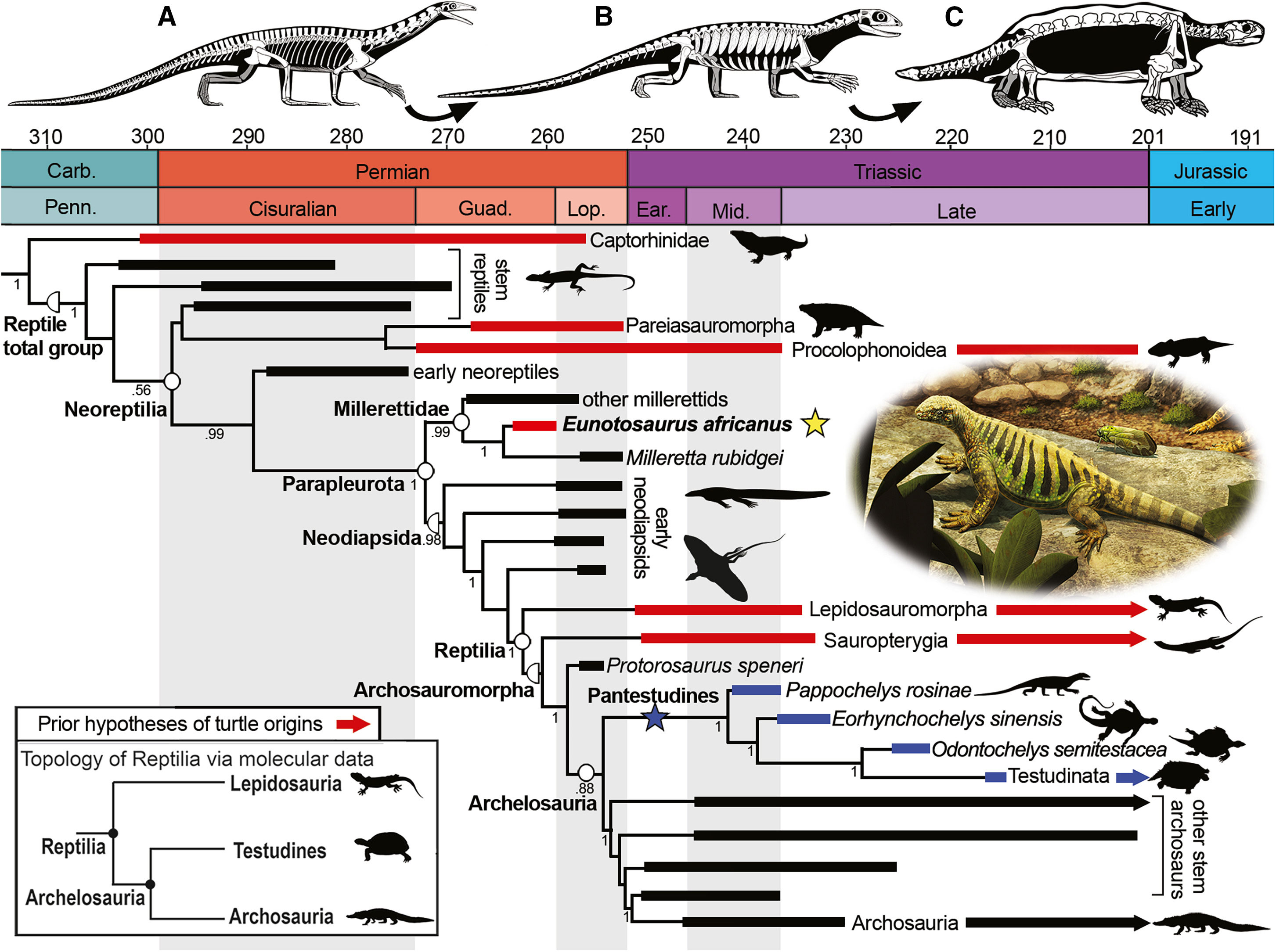
Упрощенная кладограмма, демонстрирующая родственные связи эунотозавров, архозавров и черепах. Красными линиями обозначены группы, ранее предлагавшиеся в качестве предков черепах; однозначные черепахи выделены темно-синим. Скелетные реконструкции раннего архозавроморфа проторозавра (А), раннего родственника черепах паппохелиса (B) и ранней черепахи проганохелиса (С) демонстрирует предполагаемый эволюционный переход от ранних архозавров к ранним черепахам. Рисунок из обсуждаемой статьи
Таким образом, древние родственники современных черепах оказались определенно ближе к архозаврам и другим продвинутым рептилиям, чем к примитивным миллереттидам, подобным эунотозавру. «Отрыв» эунотозавра от эволюционной линии черепах позволяет пересмотреть происхождение их панциря: поскольку наиболее ранние триасовые черепахи и их ближайшие родственники обнаружены в речных и морских отложениях, не исключено, что изначально черепаший панцирь развивался не как защита от сдавливания при рытье, а в качестве балласта для плавания. Впрочем, авторы не отрицают, что самые ранние черепашьи предки могли быть и землекопами, тем более, что они появились на Земле незадолго до массового пермского вымирания, во время которого умение рыть норы могло быть весьма полезным. Так что диверсификация и полное оформление черепах как бронированных рептилий произошло уже в триасовом периоде, одновременно с бурным расцветом их ближайших родственников-архозавроморфов, с которыми они и по сей день делят эту планету.
Источник: X. A. Jenkins, B. R. Peecook, J. N. Choiniere, V. Buffa, J. Benoit, C. Browning, V. Fernandez, K. Dollman, T. W. Gomes, G. A. McGaughey, C. J. Marchant, A. J. Fitch, M. O. Day, S. W. Evers, R. B. J. Benson. The phylogenetic origin of turtles // Current Biology. 2026. doi: 10.1016/j.cub.2026.04.070
Анна Новиковская
-
Так когда, как и почему у них закрылись оба височных отверстия? Из приведённой кладограммы видно, что черепахи исходно сидят на ветке диапсид.
-
Скорее всего, это произошло где-то между поздней пермью и средним триасом, то есть до формирования первых настоящих черепах. Основной причиной редукции височных окон, скорее всего, было увеличение прочности черепа: у черепах не может быть слишком крупная голова (иначе под панцирь не будет помещаться), поэтому нужен крепкий череп с сильной челюстной мускулатурой, чтобы прокормиться.
-
(иначе под панцирь не будет помещаться)
В итоге у изрядного количества черепах она не то, что не помещается, а принципиально сделана так, чтобы не поместиться.
Т.е. принципиально какой-нить кордилюс или кокодрил не сильно хуже защищены, чем морская или большеголовая черепахи.
И вот вопрос: на хрена было городить всю эту констрюкцию с рёбрами, сросшимися костями черепа и пр., чтобы потом от неё отказаться?
-
-


Анапсиды, диапсиды, синапсиды
-
 10.06.2026Позднепермского эунотозавра лишили звания «предка черепах»Анна Новиковская • Новости науки
10.06.2026Позднепермского эунотозавра лишили звания «предка черепах»Анна Новиковская • Новости науки
-
 13.04.2026В бассейне Кару на юге Африки впервые обнаружено яйцо синапсидыАнна Новиковская • Новости науки
13.04.2026В бассейне Кару на юге Африки впервые обнаружено яйцо синапсидыАнна Новиковская • Новости науки
-
 01.02.2025Позднетриасовый труцидоцинодон обладал не только огромными клыками, но и крупным мозгомАнна Новиковская • Новости науки
01.02.2025Позднетриасовый труцидоцинодон обладал не только огромными клыками, но и крупным мозгомАнна Новиковская • Новости науки
-
 24.12.2024Горгонопс с Майорки родился на 5 млн лет раньше своих ближайших родственников из Южной АфрикиАнна Новиковская • Новости науки
24.12.2024Горгонопс с Майорки родился на 5 млн лет раньше своих ближайших родственников из Южной АфрикиАнна Новиковская • Новости науки
-
 10.04.2024Клювастый и клыкастый рабидозаврАнна Новиковская • Картинки дня
10.04.2024Клювастый и клыкастый рабидозаврАнна Новиковская • Картинки дня
-
 30.05.2023«Российские» иностранцевии вымерли в АфрикеАнна Новиковская • Новости науки
30.05.2023«Российские» иностранцевии вымерли в АфрикеАнна Новиковская • Новости науки
-
 06.12.2022Растительноядные экзаэретодоны баловали детей более калорийной животной пищейАнна Новиковская • Новости науки
06.12.2022Растительноядные экзаэретодоны баловали детей более калорийной животной пищейАнна Новиковская • Новости науки
-
 13.10.2022Нора дииктодонаАнна Новиковская • Картинки дня
13.10.2022Нора дииктодонаАнна Новиковская • Картинки дня
-
 05.09.2022Найдены мумии листрозавров, живших в начале триасового периодаАнна Новиковская • Новости науки
05.09.2022Найдены мумии листрозавров, живших в начале триасового периодаАнна Новиковская • Новости науки
-
 29.12.2021Проколофон из кернаАнтон Ульяхин • Картинки дня
29.12.2021Проколофон из кернаАнтон Ульяхин • Картинки дня












Долгие годы позднепермский эунотозавр (две рептилии на переднем плане) считался ранним представителем эволюционной линии, ведущей к черепахам, однако новое исследование показало, что предком черепах он быть не мог. Рисунок ©Андрея Атучина с сайта eurekalert.org