В ходе эволюционного эксперимента у дрожжей появился новый ген

Использовав индуцированный мутагенез и отбор в селективной среде, ученые из Нидерландов добились того, что дрожжи вида Saccharomyces eubayanus научились сбраживать углевод мальтотриозу, которую изначально они не могли усваивать. Оказалось, что новое качество возникло за счет появления нового гена, кодирующего транспортер для мальтотриозы. Этот ген сформировался посредством сложной рекомбинации между тремя генами семейства MALT, кодирующими транспортеры другого углевода — мальтозы. Все три гена располагаются у S. eubayanus на трех разных хромосомах в субтеломерной области.
Главными героями работы, о которой пойдет речь ниже стали дрожжи. Но прежде, чем перейти к основному рассказу, поговорим немного о приятном — о пиве.
Пиво начали изготавливать многие сотни, а то и тысячи лет назад. Суть технологии в том, чтобы приготовить сначала сусло — специальное варево на основе солода (проростков семян), а затем поместить его в благоприятные условия для роста и метаболизма микроорганизмов, которые сбраживают эту массу и превращают в хмельной напиток. Сусло содержит разнообразные углеводы и другие органические соединения, которые микроорганизмы используют в качестве субстрата. Продукты метаболизма, в числе которых спирты (в первую очередь, конечно, этиловый), органические кислоты, эфиры, фенолы, и т. д., выделяются во внешнюю среду (для желающих ознакомиться с химией пива глубже — подробная таблица).
По завершении ферментации (этим термином обозначают всю совокупность химических процессов, сопровождающих процесс приготовления пива) образуется продукт очень сложного химического состава, пропорции всех компонентов которого определяют вкусовые оттенки разнообразных сортов пива, которые имеют большое значение для гурманов и знатоков этого напитка. Эти пропорции зависят от целого ряда факторов: от способа приготовления сусла (в зависимости от рецепта, в него могут добавляться шишки хмеля, травы, ягоды и т. д.), конкретных условий, при которых проходила ферментация, а также особенностей метаболизма конкретных штаммов микроорганизмов, осуществляющих сбраживание.
На заре освоения пивоварения люди, конечно, ничего не знали о том, что именно происходит в бочках во время настаивания, и благодаря чьим «стараниям» сусло превращается в хмельной напиток. Брожение происходило за счет тех микроорганизмов, которые попадали в бочки из естественной среды сами собой. Среда в бочках с суслом, однако, в наибольшей степени благоприятствует именно росту дрожжей — одноклеточных грибов (что не означает отсутствия в примеси и разнообразных бактерий). Первоначально была освоена технология, называемая сегодня верховой — ферментация при этом способе осуществляется при относительно высокой температуре — от 15°С, и длится до 6–7 дней. Пиво, получаемое таким способом, называется элем. Когда ученые занялись выделением чистых культур и классификацией микроорганизмов, выяснилось, что основным микроорганизмом, обеспечивающим сбраживание при приготовлении эля является вид дрожжей, получивший название Saccharomyces cerevisiae. Этот же грибок обеспечивает сбраживание теста для хлеба и пирожков.
В XV веке немецкие пивовары начали готовить пиво новым способом — низовым, при котором процесс брожения происходил при более низкой температуре (от 5 до 12°С) и занимал более длительное время (до 21 дня). При низовом брожении дрожжи не всплывают и не формируют пленку на поверхности сусла, как при приготовлении эля, а образуют массивный осадок на дне посуды для ферментации. Пиво, которое получается таким способом, называют лагером. По вкусу он отличается от эля, и в настоящее время преобладает на рынке пива. В 1870 г. немецкий ученый Макс Реесс выделил и описал преобладающий вид дрожжей из сусла лагерного пива, дав ему название Saccharomyces pastorianus — в честь Луи Пастера. Этот вид, как выяснилось впоследствии, более холодостойкий, чем S. cerevisiae. Вероятно, при переходе к холодному способу ферментации селекция в сторону холодостойкости произошла естественным образом.
В эру геномных исследований появилась возможность глубже изучить биологию дрожжей и разобраться в их родственных связях. Первым в 1996 г. был отсеквенирован геном S. cerevisiae. В 2000-е годы начали исследовать и описывать геномы других видов, включая как лабораторные, так и «дикие» штаммы.
Теперь можно перейти к рассказу о непосредственном герое свежего исследования — еще одном представителе дрожжей рода Saccharomyces. История этого вида весьма своеобразна — сначала его открыли заочно. Изучение генома S. pastorianus в начале XXI века привело к выводу, что это аллополиплоидный вид (встречаются триплоидные и тетраплоидные штаммы) — поскольку часть его генов соответствовала последовательности S. cerevisiae, а вторая часть явно принадлежала какому-то другому, ранее неизвестному родительскому виду (C. Monerawela, U. Bond, 2017. The hybrid genomes of Saccharomyces pastorianus: A current perspective). Интересно, что наблюдалось и некоторое количество генов «смешанного» состава, последовательность которых в одних участках соответствует S. cerevisiae, а в других — другому виду. То есть между хромосомами двух родительских видов происходила рекомбинация прямо внутри рамок считывания белок кодирующих генов.
Удивительно, но в 2011 году этот второй родительский вид, получивший свое название Saccharomyces eubayanus, был обнаружен коллективом исследователей из Португалии и Аргентины в Патогонии — южной оконечности Южной Америки (D. Libkind et al., 2011. Microbe domestication and the identification of the wild genetic stock of lager-brewing yeast)! Его обнаружили в местных лесах нотофагуса в почве под деревьями и на деревьях в ассоциации с паразитическими грибами Cyttaria hariotii (рис. 2). Опознать в S. eubayanus «прародителя» лагера помог, опять же, анализ его генома и сравнение с ранее аннотированным в международных базах геномами других дрожжей, включая S. pastorianus. Теперь уже, увы очень сложно установить, как и когда произошла судьбоносная встреча этих двух достаточно разных по экологии видов дрожжей, благодаря которой возник гибрид, и как этот гибрид оказался в Европе в бочках тех самых пивоваров-инноваторов, которые стали изобретателями лагера.

Рис. 2. Плодовые тела грибов Cyttaria hariotii на нотофагусе — вместе с ними на деревьях были обнаружены дрожжи S. eubayanus. Эти грибы производят сахаристые выделения, которые и создают благоприятную среду для роста дрожжей. Фото с сайта en.wikipedia.org
Конечно, было интересно изучить, какие именно свойства гибрид получил от каждого из родителей, и в том числе, оценить возможные перспективы использования в пивоварении нового вида S. eubayanus. Выяснилось, что этот вид, как и гибрид, отличается холодостойкостью. А что насчет метаболизма? Оказалось, что S. eubayanus отличается от S. cerevisiae и S. pastorianus полной неспособностью усваивать мальтотриозу — углевод, состоящий из трех остатков глюкозы, один из продуктов частичного расщепления крахмала. Трисахарид мальтотриоза и дисахарид мальтоза составляют основную долю углеводов в солоде. Углеводы перерабатываются дрожжами в спирт при брожении, и, соответственно, способность к их утилизации в значительной мере определяет эффективность ферментации пива. Таким образом, S. eubayanus не очень-то подходит для использования в пивоварении.
Интересно, что в 2015 году группа ученых решила воспроизвести естественным образом произошедшую гибридизацию в лаборатории (K. Krogerus et al., 2015. New lager yeast strains generated by interspecific hybridization). Они поместили в общий сосуд S. cerevisiae и S. eubayanus и успешно получили гибрид, который унаследовал от обоих родителей все самое лучшее: холодостойкость и способность сбраживать сусло при низких температурах от S. eubayanus и эффективное усвоение разнообразных углеводов от S. cerevisiae. Более того, этот штамм позволял получить более крепкое пиво (то есть с более высоким содержанием спирта), чем оба родительских!
Авторами нового исследования, о котором пойдет речь ниже, стал коллектив ученых из Нидерландов под руководством Жан-Марка Дарана (Jean-Marc Daran). Его лаборатория специализируется на изучении биологии дрожжей с 2001 года, являясь одним из лидеров в данной области (ежегодно ее сотрудники публикуют более десятка работ). Они решили попробовать «научить» S. eubayanus усваивать мальтотриозу. Ведь вполне может оказаться, что таким образом будет получен новый ценный штамм, который будет обладать какими-то своими преимуществами и пригодится в пищевой промышленности.
Ученые решили применить для решения поставленной задачи подход «эволюции в пробирке». Это вполне типичная задача для селекционеров, работающих, к примеру, над улучшением пород животных или сортов растений. К тому же у авторов был уже достаточно большой и успешный опыт по применению такого подхода для улучшения хозяйственно-ценных признаков у дрожжей: ранее они опубликовали несколько работ с описанием разработанной ими методологии и полученных результатов (вот три примера: первый, второй и третий). Аналогичные работы в последнее время успешно выполнялись и другими коллективами. Ученым удавалось повысить усвоение тех или иных углеводов, увеличить выход определенных продуктов метаболизма, улучшающих вкусовые качества продукта, снизить чувствительность дрожжей к высоким температурам или присутствию в среде химических ингибиторов некоторых важных дрожжевых ферментов.
Эволюционный подход подразумевает, что организмам создаются, во-первых, условия для генерирования генетического разнообразия (этот процесс можно стимулировать разными мутагенными воздействиями — например, ультрафиолетом, химическими мутагенами и т. д., — которые вносят повреждения в ДНК, что, в свою очередь активирует в клетках процессы репарации), а во-вторых, условия для селекции — благоприятствующие выживанию носителей желательных для исследователя особенностей фенотипа. Конечно, можно было бы пойти и по более прямому пути генной модификации (этой технологией авторы тоже отлично владеют) — просто ввести в геном S. eubayanus недостающие гены для усвоения мальтотриозы. Но в «эволюционном» подходе есть свои важные преимущества: во-первых, так можно получить хороший результат, реализуемый непредсказуемым способом, который может оказаться лучше, чем при использовании просчитанного человеком «генетического» решения. А во-вторых, и для ученых это не менее важно, — такой подход позволяет одновременно больше узнать об ассоциации тех или иных белков и путей с их функциями и с фенотипическими характеристиками конкретного организма. То есть таким образом совмещается эффективное решение как практических, так и фундаментальных исследовательских задач.
Прежде всего, нужно было разобраться, как вообще генетически обеспечена утилизация углеводов типа мальтозы и мальтотриозы у дрожжей, в частности у S. eubayanus. Все дрожжи имеют в своих хромосомах по несколько специальных кластеров MAL, каждый из которых, как правило, включает три гена: MALT — ген белка-транспортера, специфичного к тому или иному типу углевода, MALS — ген белка-фермента для расщепления углеводов и MALR — ген белка-регулятора, управляющего экспрессией первых двух (а иногда и самого себя). Сам ген-регулятор активируется в ответ на повышение концентрации в среде «своего» углевода (рис. 3). Эти кластеры паралогичны (родственны) друг другу и отвечают за утилизацию мальтозы. Транспортер мальтотриозы, имеющийся, к примеру, у S. cerevisiae, кодируется другим неродственными геном, AGT1. У S. eubayanus функциональных транспортеров мальтотриозы до недавнего времени не обнаруживалось (хотя об одном исключении ученые недавно все же сообщили, см.: E. P. Baker et al., 2019. Evolution of a novel chimeric maltotriose transporter in Saccharomyces eubayanus from parent proteins unable to perform this function).

Рис. 3. Организация системы утилизации мальтозы и мальтотриозы у представителей рода Saccharomyces. Вверху — типичная организация кластера, состоящего из генов транспортера углевода (MALT), фермента, обеспечивающего его расщепление (MALS — это фермент, способный расщеплять как мальтозу, так и мальтотриозу) и регуляцию экспрессии генов в кластере. Cиним цветом показаны известные кластеры, связанные с утилизацией мальтозы (MAL, MPH) и ген транспортера мальтотриозы (AGT1) у S. cerevisiae, оранжевым — четыре гена транспортера в кластерах MAL в геноме S. eubayanus. Римскими цифрами подписаны номера хромосом. Возле хромосомы с геном AGT1 в геноме S. eubayanus стоит знак вопроса, поскольку в литературе имеется единичное упоминание о присутствии этого гена в геноме данного вида (E. P. Baker et al., 2019. Evolution of a novel chimeric maltotriose transporter in Saccharomyces eubayanus from parent proteins unable to perform this function), а у штамма, с которым работали авторы обсуждаемого исследования, такой ген не был обнаружен. Рисунок из обсуждаемой статьи в PLOS Genetics
У S. eubayanus, как выяснилось, таких кластера четыре: на хромосомах II, V, XIII и XVI. Все четыре кластера располагаются в субтеломерных (приконцевых) участках хромосом. Гены белков-транспортеров в этих кластерах обозначили, соответственно MALT1, MALT2, MALT3 и MALT4. При проверке их функциональности, которая предшествовала обсуждаемой работе (A. Brickwedde et al., 2018. Structural, Physiological and Regulatory Analysis of Maltose Transporter Genes in Saccharomyces eubayanus CBS 12357T), ученые из группы Дарана выяснили следующее:
1) Гены MALT1 и MALT3 почти не экспрессируются (даже в присутствии мальтозы), предположительно вследствие того, что в соответствующих кластерах отсутствуют необходимые регуляторные последовательности. Таким образом, они практически не играют заметной роли в утилизации углеводов дрожжами.
2) MALT2 и MALT4 активно экспрессируются в присутствии мальтозы, обеспечивая эффективный транспорт в клетки мальтозы, но не мальтотриозы.
Чтобы добиться формирования у дрожжей нужного свойства, нужно было заставить их выживать на селективной среде. В данном случае использовали питательный бульон, в котором не было никакого другого источника питательных веществ, кроме мальтотриозы. Дело в том, что нельзя было исключить, что хотя бы некоторые клетки среди S. eubayanus на самом деле способны (пусть слабо) поглощать мальтотриозу, но ее усвоение подавляется в присутствии других углеводов (такая взаиморегуляция путей утилизации углеводов достаточно характерна для дрожжей, в том числе и S. pastorianus). Три месяца испытатели ждали, что появятся признаки роста дрожжей, способных размножаться на этой среде, но так ничего и не дождались.
Тогда ученые решили подстегнуть процесс и подвергли дрожжевые клетки воздействию ультрафиолета. Дозу рассчитали так, чтобы выживала половина всех клеток. Эта обработка резко повысила генетическое разнообразие среди клеток дрожжей, и теперь можно было переходить, собственно, к этапу селекции. Дрожжи снова заставили выживать на той же среде с мальтотриозой в качестве единственного источника углеводов. Стратегия сработала. Через две недели стал заметен рост «выживанцев», а еще через три недели зафиксировали уменьшение содержания мальтотриозы в среде до половины от исходной концентрации.
Далее высеяли клетки на твердую (агаризованную) среду все с той же мальтотриозой и получили несколько отдельных клонов, с которыми продолжили работу. Конечной целью было получить штаммы, годные для пивоваренной промышленности, так что теперь стали растить дрожжи на сусле. Тут выяснилось, что полученные мутанты не желают усваивать мальтотриозу в условиях, где им в достатке предоставлены другие углеводы. Тогда ученые стали растить их на разбавленном (в 6 раз) сусле, обогащенном мальтотриозой. Это сработало. Спустя 121 день (авторы указывают, что это соответствует примерно 125 поколениям) дрожжи уже без проблем усваивали все компоненты сусла не хуже, чем традиционные штаммы, используемые в пивоварении. При этом они нарабатывали больше спирта, чем исходный штамм (рис. 4).
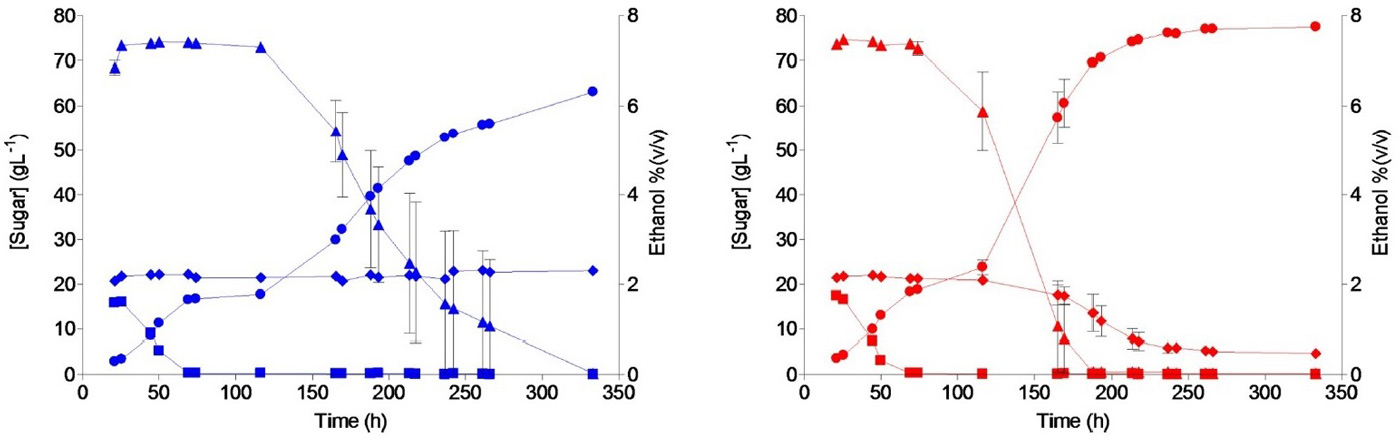
Рис. 4. Изменение концентрации углеводов и спирта при выращивании дрожжей на сусле в биореакторе. Левый график — исходный штамм, правый — штамм полученный по завершении всех этапов селекции. Линии с маркерами в виде треугольников, ромбов и квадратов отражают, соответственно, изменения концентрации мальтозы, мальтотриозы и глюкозы (значения указаны по левой вертикальной оси), линии с круглыми маркерами отражают накопление спирта (правая вертикальная ось). Рисунок из обсуждаемой статьи в PLOS Genetics
Теперь оставалось выяснить, что же именно произошло на уровне генов, откуда взялся новый транспортер. Заметим, что в ходе селекции было три ключевых этапа: до начала селекции (контроль), затем — штаммы, полученные после мутагенеза и первичной селекции на искусственной среде с мальтотриозой, и, наконец, — штаммы, полученные в итоге после селекции на способность усваивать мальтотриозу из сусла.
Выяснилось, что штаммы первого этапа селекции отличались от контрольных некоторым количеством однонуклеотидных полиморфизмов (SNP), а также полиморфизмов типа инсерций и делеций, из которых ни один не попал в области MAL-кластеров. Как эти мутации помогли дрожжам выживать на мальтотриозе и хоть медленно, но усваивать ее, — осталось, увы, совершенно неясным. Но вот штаммы, полученные на конечном этапе, преподнесли хороший сюрприз: у них ген MALT4, расположенный на XVI хромосоме, оказался изменен и представлял собой мозаику из последовательностей, происходящих из генов MALT1, MALT3 и MALT4. То есть выходит, произошла сложная рекомбинация с участием трех хромосом, в результате которой получился новый ген (рис. 5)! И он действительно выполнял новую функцию: теперь это был уже транспортер мальтотриозы, но не мальтозы. В качестве транспортера мальтозы остался ген MALT2.

Рис. 5. Изменения в геномах штаммов, полученных при селекции S. eubayanus. А — количество мутаций разных типов, зафиксировавшихся у штаммов, которые научились утилизировать мальтотриозу после первого (верхний круг) и после завершающего (нижний круг) этапов селекции. SNP — однонуклеотидные полиморфизмы, INDEL — мутации типа небольших инсерций или делеций, CNV — мутации типа изменения числа копий обширных сегментов хромосом. Мутации, породившие новый ген, входят в число CNV, поскольку части генов MALT1 и MALT3 оказались продублированы на XVI хромосоме в составе гена MALT413. B — схема концевого участка левого плеча XVI хромосомы у мутантного штамма, полученного после селекции. Показано две одинаковых хромосомы, поскольку дрожжи имеют диплоидный геном. Разными цветами обозначены и подписаны сегменты двух других хромосом, внедрившиеся в XVI хромосому в результате рекомбинации и частичной конверсии. Можно заметить, что имело место не одно, а как минимум 3 (если не 4) рекомбинационных события. С — мозаичная структура гена MALT413, включающая в себя участки от генов MALT1, MALT3 и MALT4 (показаны все гены, числа указывают на степень сходства нуклеотидных последовательностей их сегментов с последовательностью новообразованного гена). D — биоинформатически предсказанная структура белка MALT413 при виде «в профиль» и сверху (при открытом состоянии канала). Рисунок из обсуждаемой статьи в PLOS Genetics
Убедиться в том, что определяющим в обеспечении нового признака является именно этот новый ген (его обозначили MALT413) позволили два дополнительных эксперимента. Первый эксперимент — введение этого гена в исходный штамм — дрожжи сразу приобретали способность роста на среде с мальтотриозой и усвоения мальтотриозы из сусла (хотя и более слабую, чем у штамма после селекции — что говорит о том, что вспомогательную роль в утилизации мальтотриозы играют и еще какие-то дополнительные мутации). Второй — обратный: в полученном в результате селекции штамме нокаутировали ген MALT413 — и эти дрожжи сразу переставали усваивать мальтотриозу из сусла, а на мальтотриозной среде росли гораздо хуже. Между тем, введение в клетки дополнительных копий транспортера мальтозы MALT2 на усвоение мальтотриозы никак не влияло (рис. 6). Нокаут же MALT2 в штамме, прошедшем селекцию, приводил к утрате способности клеток утилизировать мальтозу. И инсерцию генов в исходном диком штамме, и нокаут генов в селектированном штамме осуществляли с использованием CRISPR/Cas9, ставшего уже почти неотъемлемым в арсенале молекулярных биологов.

Рис. 6. Проверка функциональности генов-транспортеров у S. eubayanus. A — схема осуществляемой инсерции гена-транспортера в геном дрожжей при помощи системы CRISPR/Cas. Ген встраивали на место другого дрожжевого гена, SGA1, который не был нужен дрожжам в условиях эксперимента (в норме этот ген участвует в процессе споруляции, и не функционирует в фазе активного роста). Это место в геноме дрожжей было подобрано экспериментаторами в прошлые годы как вполне удобное с точки зрения подобных задач: гены стабильно работают, и при этом модификация не нарушает никаких других жизненно важных функций клеток. На графиках показана динамика утилизации углевода экспериментальными штаммами S. eubayanus на искусственных средах с разными типами сахаров (B — среда с глюкозой, C — с мальтозой, D — с мальтотриозой). На каждом графике синяя линия с квадратными маркерами соответствует штамму дикого типа; голубая линия (с маркерами в виде перевернутого треугольника) — штамму с искусственно введенной копией гена MALT2; фиолетовая линия (с маркерами в виде ромбов) — штамму с искусственно введенной копией гена MALT413; красная линия с треугольными маркерами — штамму, полученному по завершении всех этапов селекции на способность утилизации мальтотриозы. Рисунок из обсуждаемой статьи в PLOS Genetics
Так, в очередной раз мы видим, как эволюционные механизмы оказываются на службе у людей, позволяя получать организмы с полезными свойствами. Но не менее важна и фундаментальная составляющая этого исследования — оно демонстрирует один из путей, которыми «располагает» природа для получения новых функциональных генов. Конечно, такое быстрое развитие событий достижимо только при условии достаточно высокой интенсивности мутагенеза, отбора и размножения. В данном конкретном случае подыграло и еще несколько специфических факторов. Во-первых, это то, что у S. eubayanus уже имелось несколько генов-транспортеров химически схожих углеводов, так что новый ген мог быть собран посредством комбинирования частей этих «заготовок» (конечно, это происходило вслепую, без участия чьего-либо намерения — удачный вариант сформировался случайно, но неслучайно оказался отобран в итоге). Во-вторых, то, что гены располагались в субтеломерных областях. В этих участках хромосом всегда имеется повышенное содержание повторов, что предрасполагает к увеличению частоты рекомбинации. Третьим благоприятным фактором было наличие «запасной» копии транспортера мальтозы. Это позволило дрожжам приобрести новую полезную функцию, не потеряв уже имеющуюся. Еще одна из удивительных вещей, выявленных в этой работе — полученный ген MALT413 удивительно схож по структуре с геном транспортера мальтотриозы MTY1, обнаруженным у гибридного вида S. pastorianus. Весьма вероятно, здесь представлен случай параллельной молекулярной эволюции (возникновение схожих признаков за счет схожих мутаций).
Источник: Nick Brouwers, Arthur R. Gorter de Vries, Marcel van den Broek, Susan M. Weening, Tom D. Elink Schuurman, Niels G. A. Kuijpers, Jack T. Pronk, Jean-Marc G. Daran. In vivo recombination of Saccharomyces eubayanus maltose-transporter genes yields a chimeric transporter that enables maltotriose fermentation // PLOS Genetics. 2019. DOI: 10.1371/journal.pgen.1007853.
См. также:
1) Найден ген, превращающий простые листья в сложные, «Элементы», 18.02.2014.
2) Ген, работающий в мышцах и костях, у обезьян стал регулировать развитие мозга, «Элементы», 14.11.2016.
3) Процесс появления новых ферментов прослежен в эволюционном эксперименте, «Элементы», 23.10.2012.
4) Ученые открыли, как архебактерии выживают на уксусе, «Элементы», 25.01.2011.
5) В долгосрочном эксперименте зафиксировано поэтапное формирование эволюционного новшества, «Элементы», 25.09.2012.
Татьяна Романовская


Ну а мне до здешних корифеев духа огромноеспасибозастатью еще далековато, поэтому у меня вопрос: как облучение привело к множественной рекомбинацией в одном (единственном), отдельно взятом, гене? Какой механизм между этими явлениями?
Со снипами все ясно, но множественная рекомбинация тут каким боком вылезла?
-
Эцих, спасибо за Вашу любознательность! Вы совершенно правы, тут много чего остается неясным в смысле механизма. На самом деле, выглядит почти невероятным, что нужная комбинация мутаций в таком узком месте нашлась за такое короткое время, собственно, это и побудило рассказать здесь об этом замечательном случае.
Но вот некоторые "спекуляции", которые приходят в голову по этому поводу.
Во-первых, была все же и еще одна рекомбинация - перенос теломерного участка хромосомы 8 на хромосому 2, смысл которой для приобретения нужного свойства остался в работе нераскрытым (может его и нет, просто прошла через селекцию вместе с другими полезными мутациями по механизму автостопа). На рис 5A описаны все обнаруженные мутации, и с рекомбинациями там связаны мутации типа CNV, - всего их 5, 4 из которых поучаствовали в формировании гена MALT413. Логика подсказывала, что новый транспортер если и появится, то скорее всего на основе уже имеющихся, поэтому в первую очередь на этот ген авторы и обратили внимание. К тому же именно эти рекомбинации определили получение успешных штаммов второго этапа селекции.
Во-вторых, гомологичная рекомбинация - достаточно обычное явление в клетках, так как это один из способов восстановления серьезных повреждений ДНК. Появление таких повреждений может быть связано с внешними воздействиями (типа, например, УФ), а может происходить и спонтанно по внутренним причинам. [Чем конкретно они были вызваны в этом эксперименте - остается неясным, ведь УФ применили только в самом начале, а все "интересные" рекомбинации появились уже на втором этапе, когда просто вели селекцию]. Правда "законная" гомологичная рекомбинация должна происходить между парой гомологичных хромосом. Наши 4 MALT гена - паралоги, то есть это гомологичные гены, располагающиеся на разных (негомологичных) хромосомах. При достаточно высокой степени сходства таких паралогов система гомологичной рекомбинации может "по ошибке" совершить "незаконную" рекомбинацию (то, что такое нередко случается - известно уже давно из работ по молекулярной биологии), и тогда мы будем наблюдать то, что наблюдалось в данном случае. Кстати, о том, что гомологичная рекомбинация очень активно работала в клетках в процессе эксперимента можно заключить по повышенной гомозиготности штаммов, полученных после селекции (по сравнению с исходными). Повышение гомозиготности говорит опять же о том, что во многих местах происходили рекомбинации с конверсиями гомологичных участков, так что сайты, которые были гетерозиготными (разными на гомологичных хромосомах) теперь превращались в гомозиготные (одинаковые).
Действительно необычно здесь то, что таких рекомбинаций произошло сразу несколько на одном узком участке и еще то, что рекомбинация поменяла последовательность только в одном из 4-х генов, а в трех других хромосомах последовательность не нарушилась (включая даже те две,с которыми, собственно, происходила рекомбинация). Такое принципиально возможно, если при рекомбинации происходил не взаимный обмен участками хромосом, а "генная конверсия". Сложно здесь объяснить подробно, но есть статья с таким названием на википедии. Вкратце, смысл в том, что фрагмент одной хромосомы копируется по образцу другой. Типа как, когда задали сочинение, слабый ученик может взять тетрадку у умного однокласника, добавить в свое сочинение некоторые абзацы из сочинения товарища, и вернуть тетрадку обратно - теперь изменения произошли в одной тетради, но не в другой. Теперь учитель при проверке сочинений встретит одинаковый текст дважды - вот и получается мутация типа CNV - вариация числа копий, конкретнее, дупликация. Возвращаясь к эксперименту, выходит, что все рекомбинации меняли всегда только один ген. Но не забывайте, что речь идет о селекции. И значит, дело было не так, что случались обязательно только такие варианты мутации, а так, что среди множества мутантов, именно такой вариант оказался "выживанцем". Если бы они на завершающем этапе оставили больше вариантов полученных клонов, то, скорее всего (я предполагаю), нашлись бы и такие, у которых рекомбинация затронула и другие гены. Ну и, как уже сказано в самом синопсисе, расположение в субтеломерных областях дополнительно предрасполагает к событиям гомологичной рекомбинации. Такие соображения.
А с другой стороны, может есть какая-то принципиальная причина, почему конверсия шла бы всегда (или преимущественно) именно в 16-ю хромосому, а не в 3 остальные. Я имею в виду какие-то причины, привязанные к каким-то молекулярным механизмам, которые еще предстоит выяснить. Тоже вполне допускаю.-
Спасибо за объяснения, здесь есть над чем почесать репу.
Все же, я полагаю, последовательная, множественная рекомбинация в одном гене есть событие невероятное, теория вероятности не пропустит такое событие. Скорее, облучение привело к мутации в этом гене изначально в делящихся клетках с последующей рекомбинацией между хромосомами дочерней и материнской клетки, то есть изначально возникла популяция из организмов, гетерогенных по этому признаку. А может быть (я не дрожжевик) потом произошла еще и коньюгация между генетически отличными клетками с рекомбинацией и дальше уже отбор завершил дело.
Загадка интересная.
-
Вот человека создали по образу и подобию. И он теперь творит. Облучает ультрафиолетом, индуцирует мутации, создаёт среду. А в итоге новый ген. Но это же зделал Всевышний. Только и намеренно облучать не приходилось. Всё зделал большой взрыв.
полученный ген MALT413 удивительно схож по структуре с геном транспортера мальтотриозы MTY1, обнаруженным у гибридного вида S. pastorianus.Чисто теоретически. А может появиться группа ученных, которые геноинженерными методами сконструируют новый "штамм", а потом нам расскажут что-нибудь о сложных экспериментах?
Эволюционные эксперименты
-
 17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки
17.03.2025Полногеномная дупликация дает немедленный адаптационный выигрышЕлена Наймарк • Новости науки -
 14.01.2022Замена небольшого участка генома на синонимичный заставляет бактерию иначе решать ту же эволюционную задачуТатьяна Романовская • Новости науки
14.01.2022Замена небольшого участка генома на синонимичный заставляет бактерию иначе решать ту же эволюционную задачуТатьяна Романовская • Новости науки
-
 14.01.2021В ходе эволюционного эксперимента удалось получить генералистов при симпатрическом видообразованииЕлена Наймарк • Новости науки
14.01.2021В ходе эволюционного эксперимента удалось получить генералистов при симпатрическом видообразованииЕлена Наймарк • Новости науки
-
 18.11.2019Новый метод генетического штрихкодирования позволяет детально следить за эволюцией дрожжейАлександр Марков • Новости науки
18.11.2019Новый метод генетического штрихкодирования позволяет детально следить за эволюцией дрожжейАлександр Марков • Новости науки
-
 11.11.2019Симбиотические дрожжи помогают дрозофилам адаптироваться к соленому кормуАлександр Марков • Новости науки
11.11.2019Симбиотические дрожжи помогают дрозофилам адаптироваться к соленому кормуАлександр Марков • Новости науки
-
 15.08.2019В ходе эволюционного эксперимента у дрожжей появился новый генТатьяна Романовская • Новости науки
15.08.2019В ходе эволюционного эксперимента у дрожжей появился новый генТатьяна Романовская • Новости науки
-
 06.11.2018В ходе эволюционного эксперимента патогенный гриб превратился в полезного симбионтаАлександр Марков • Новости науки
06.11.2018В ходе эволюционного эксперимента патогенный гриб превратился в полезного симбионтаАлександр Марков • Новости науки
-
 16.10.2018Способствует ли адаптация к разным диетам развитию репродуктивной изоляции?Александр Марков • Новости науки
16.10.2018Способствует ли адаптация к разным диетам развитию репродуктивной изоляции?Александр Марков • Новости науки
-
 14.10.2018Успех адаптации мух Drosophila melanogaster к избытку соли в корме зависит от дрожжей определенного видаСергей Ивницкий • Журнал общей биологии • №5, 2018
14.10.2018Успех адаптации мух Drosophila melanogaster к избытку соли в корме зависит от дрожжей определенного видаСергей Ивницкий • Журнал общей биологии • №5, 2018
-
 06.03.2018Мухи, приспособленные к неоднородной среде, получают эволюционное преимуществоЕ. Яковлева, А. Горшкова, Е. Фетисова • Журнал общей биологии • №1, 2018
06.03.2018Мухи, приспособленные к неоднородной среде, получают эволюционное преимуществоЕ. Яковлева, А. Горшкова, Е. Фетисова • Журнал общей биологии • №1, 2018
Новости: Эволюция


См. также








Рис. 1. Saccharomyces cerevisiae — наиболее известный вид дрожжей: с их помощью производятся хлеб, пиво и вино. Всего описано около 30 видов рода Saccharomyces, населяющих самые разные уголки земного шара. Внешне они малоотличимы друг от друга. По оценкам ученых, основатель рода жил 10–20 млн лет назад. На этой микрофотографии каждый «шарик» — это одна дрожжевая клетка размером 5–10 мкм (что в 20 раз меньше толщины человеческого волоса). Можно заметить почкующиеся клетки. Фото с сайта pixels.com