Эволюция муравьев на «небесных островах» Аризоны оказалась отчасти предсказуемой
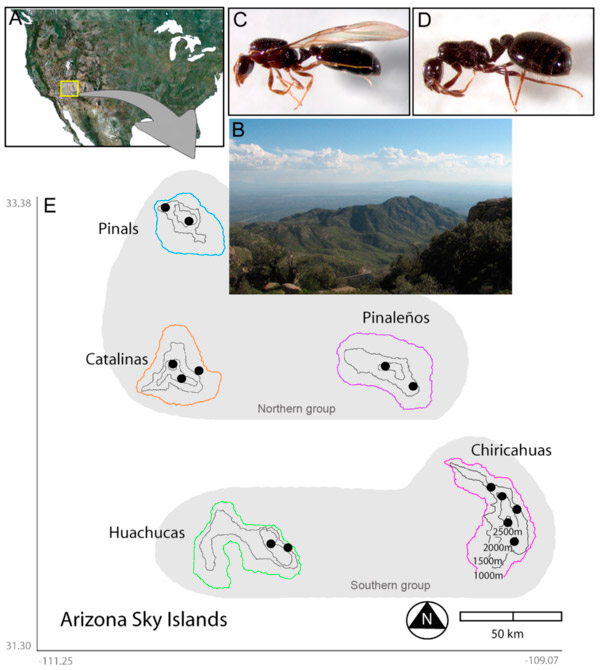
Пять популяций муравьев Monomorium emersoni, обитающих на пяти разделенных пустыней горах («небесных островах») в Южной Аризоне, независимо друг от друга приобрели важный новый признак — бескрылых самок. Как выяснилось, в основе этого эволюционного новшества лежали по большей части схожие, но всё же немного различающиеся изменения экспрессии регуляторных генов в зачатках крыльев. Сходство эволюционных изменений, по-видимому, связано не только с одинаковой направленностью отбора, но и со сходными эволюционными возможностями популяций, которые, в свою очередь, определяются единой генетической основой регуляции индивидуального развития. Различия отражают предшествующую историю популяций и предположительно связаны с разными наборами нейтральных мутаций, накопленных ими за время изолированного существования.
Давно известно, что уединенные острова являются своеобразными эволюционными лабораториями, где животные и растения могут претерпевать весьма быстрые и причудливые изменения. Этому способствует целый ряд факторов. Важнейшими из них, по-видимому, являются изоляция (она препятствует скрещиванию с чужаками и «размыванию» возникающих отличий), малый размер популяций и эффект основателя (это повышает вероятность быстрых изменений за счет генетического дрейфа), а также пониженная межвидовая конкуренция (обилие свободных ниш в ненасыщенных экосистемах).
Всё это характерно не только для настоящих островов, но и для других разобщенных, фрагментированных местообитаний. Например, изолированные озера могут выступать в роли «островов» для пресноводных рыб, а разобщенные горные массивы — для высокогорной флоры и фауны (см.: Теория островной биогеографии).
Такие отдельно стоящие горные массивы, природные условия которых резко контрастируют с окружающей их равниной, называют «небесными островами» (sky islands). Самый известный из таких массивов — это Мадреанские «небесные острова» (Madrean Sky Islands) — изолированные горы в штатах Нью-Мексико и Аризона (США) и штатах Чиуауа и Сонора (Мексика).
Авторы обсуждаемой статьи исследовали фауну той части Мадреанских «небесных островов», которая находится в штате Аризона. Это группа из пяти гор на юге Аризоны, разделенных пустынными равнинами. Обитатели гор, не способные пересечь пустыню, фактически находятся в изоляции, как на настоящем острове. Подобно другим уединенным архипелагам, «небесные острова» являются действующей «эволюционной лабораторией». Об этом свидетельствуют, в частности, имеющиеся там эндемичные (больше нигде не встречающиеся) виды растений, улиток, муравьев и грызунов.
Палеонтологические данные позволили детально реконструировать историю этого уникального «архипелага». Во время последнего оледенения, 90–20 тысяч лет назад, климат в регионе был намного прохладнее, и на разделяющих горы равнинах росли леса. Сами горы были тогда малопривлекательным местом: там было очень холодно. 20–10 тысяч лет назад в регионе происходило быстрое потепление, которое в итоге привело к опустыниванию равнин. Леса вместе со своими обитателями постепенно отступали вверх, на горные склоны, где было прохладнее и выпадало много осадков. В настоящее время на склонах аризонских «небесных островов» существуют очень резкие экологические градиенты — возможно, самые резкие в Северной Америке. На малых высотах, где относительно тепло и сухо, растут дубово-можжевеловые леса. В высокогорьях, где влажно и холодно, преобладают хвойные. Перемещение всего на несколько сотен метров вверх по склону здесь соответствует, с экологической точки зрения, путешествию на сотни или даже тысячи километров на север по равнине.
Один из обитателей «небесных островов» Аризоны, муравей Monomorium emersoni, живет на всех пяти горах. Он встречается на разных высотах, причем в нижней части склонов пригодные для него местообитания, как правило, менее фрагментированы, чем в высокогорьях.
Группа биологов из Канады, США и Австрии провела всесторонний анализ морфологии, генетики и индивидуального развития этих муравьев, что позволило понять их популяционную историю и пути адаптации к необычным местным условиям. Особенностью исследования является его многогранность: для решения поставленных задач авторы привлекли данные и методы самых разнообразных дисциплин, включая палеонтологию, экологию, генетику, биоинформатику и эволюционную биологию развития.
Судя по распределению генетического полиморфизма (вариантов митохондриальных и ядерных генов), муравьи, обитающие ныне на пяти «небесных островах», 100–200 тысяч лет назад представляли собой единую популяцию. Около 80 тысяч лет назад они подразделились на две части, от одной из которых произошли нынешние обитатели трех северных гор, а от другой — двух южных. Во время потепления, произошедшего 20–10 тысяч лет назад, муравьиное население каждой из гор оказалось в почти полной изоляции, которая сохраняется по сей день. В настоящее время генетического обмена между пятью популяциями практически нет.
Авторы обнаружили, что муравьи успешно приспособились к локальным температурным условиям. Семьи, обитающие в холодных местах, лучше переносят холод (быстрее приходят в себя после охлаждения), а жители жарких нижних частей склонов отличаются повышенной устойчивостью к перегреву. При этом тепло- и холодолюбивые обитатели одной и той же горы более родственны друг другу, чем жителям других гор, обладающим сходными температурными предпочтениями. Это значит, что адаптация к местным температурным условиям, скорее всего, шла параллельно на всех пяти горах.
Кроме того, авторы обнаружили, что местные популяции отличаются друг от друга по частоте встречаемости бескрылых самок (рис. 1). Здесь нужно пояснить, что наличие или отсутствие крыльев у самок — важный признак, от которого зависят расселительные возможности вида. Обычно у муравьев функция расселения возложена на крылатых самок, которые после спаривания с крылатыми самцами основывают новые семьи в одиночку, без помощи рабочих из родительского гнезда. Однако во многих эволюционных линиях параллельно и независимо появился другой, менее рискованный способ основания новых семей — при помощи бескрылых самок, которые спариваются с самцами в родительском гнезде и отправляются на поиски нового места жительства пешком, в сопровождении группы рабочих. Конечно, утрата способности к полету отрицательно сказывается на расселительных возможностях, зато резко снижает смертность самок в процессе расселения. Эволюционный переход от первого ко второму способу расселения может быть поддержан отбором, если шансы найти подходящее место рядом с родным гнездом велики, а дальние перелеты крайне рискованны.
На каждом из пяти аризонских «небесных островов» были обнаружены как крылатые, так и бескрылые самки-основательницы (речь идет, конечно, о молодых самках, которые покидают родное гнездо либо по воздуху, либо пешком, а не о тех, кто уже приступил к строительству нового гнезда, избавившись от ненужных теперь крыльев). Каждая муравьиная семья производит либо только крылатых самок, либо только бескрылых, либо тех и других. Выбор будущими самками одного из двух альтернативных фенотипов в ходе развития контролируется как генетическими, так и средовыми факторами.
Авторы сопоставили частоту встречаемости бескрылых самок с локальными условиями и пришли к выводу, что из всех возможных факторов, которые в принципе могли бы способствовать отбору на бескрылость, актуальными являются два: высота над уровнем моря и фрагментированность местообитаний. Чем выше по склону живет муравьиная семья и чем сильнее фрагментированы в данной местности пригодные для жизни муравьев местообитания, тем выше вероятность, что самки, производимые данной семьей, окажутся бескрылыми. Как уже говорилось, фрагментарность местообитаний в целом растет с высотой, но эта корреляция не абсолютна, и по отдельности каждый из двух факторов хуже объясняет распределение бескрылых самок, чем оба фактора вместе. Другие факторы, которые тоже могли бы влиять на выгодность бескрылости, такие как температурные условия и продуктивность биотопа (обилие пищи), не коррелируют с частотой встречаемости бескрылых самок.
Судя по совокупности генетических данных, для предковой популяции муравьев с «небесных островов» Аризоны были характерны крылатые самки. Отчасти это отражается в их распределении по склонам: крылатые самки преобладают в нижних частях склонов, которые были заселены муравьями раньше. Бескрылые формы появились параллельно и независимо на каждой из пяти гор уже после того, как популяции оказались отделены друг от друга пустынными равнинами, а на вершинах сформировались нынешние фрагментированные местообитания (где преобладают бескрылые самки). Параллельное появление бескрылых самок подтверждается, в частности, тем, что бескрылые самки генетически ближе к крылатым самкам с той же горы, чем к бескрылым самкам с других гор. Генетическая вариабельность изученных муравьев в целом намного лучше коррелирует с географией (и отражает историю разделения исходной популяции сначала на две, а потом на пять частей, соответствующих разным горам), чем с местными условиями среды и фенотипом (крылатостью или бескрылостью самок). Это и понятно, ведь большая часть генетической вариабельности — это нейтральные различия, не подконтрольные отбору и не связанные с адаптивными признаками.
Для параллельных эволюционных изменений недостаточно одной лишь сходной направленности отбора: нужно еще, чтобы в разобщенных популяциях, подвергающихся такому отбору, с достаточной частотой появлялись сходные наследственные вариации, с которыми отбор мог бы работать. Спектр наследственной изменчивости, в свою очередь, определяется структурой регуляторных генных сетей, управляющих развитием. Эти генные сети, как известно, обладают очень высоким эволюционным консерватизмом, то есть медленно меняются в ходе эволюции и часто оказываются похожими даже у очень далеких друг от друга животных. Отсюда — такие удивительные случаи гомологичной наследственной изменчивости, как, например, аномалии в развитии глаз, возникающие у насекомых и млекопитающих в результате мутаций одного и того же гена (см. PAX6). Это вполне естественно: сходные регуляторные системы допускают сходные наборы возможных модификаций или поломок, порождающих сходные изменения фенотипа (см.: Гомологические ряды в наследственной изменчивости). Именно этим сходством в значительной мере объясняется направленный и отчасти предсказуемый характер эволюции.
У муравьев, похоже, есть сильная предрасположенность к формированию бескрылых самок, ведь это эволюционное событие происходило у них независимо около 200 раз. Из 16 видов рода Monomorium, встречающихся в Северной Америке, у шести известны только крылатые самки; у пяти других — только бескрылые; наконец, у оставшихся пяти видов (в том числе у M. emersoni) наряду с крылатыми самками встречаются также и бескрылые: у кого-то — как редкая аномалия, у кого-то — как альтернативный вариант нормального фенотипа. Таким образом, появление бескрылых самок у муравьев вообще и у Monomorium в частности — высоковероятное эволюционное событие. Возможно, такая предрасположенность связана с тем, что муравьи с самого начала своей эволюционной истории умели на основе одного и того же генотипа производить как крылатых особей (цариц), так и бескрылых (рабочих). При этом выбор пути развития определяется внешними, негенетическими факторами, такими как температурный режим и диета личинки. Иными словами, уже у первых муравьев структура регуляторных генных сетей позволяла блокировать программу развития крыльев в том случае, если развивающаяся особь собиралась стать рабочей, а не царицей. Чтобы сделать на этой основе бескрылую самку, нужно просто скопировать механизм блокировки развития крыльев из алгоритма развития рабочей особи в алгоритм развития самки.
Исходя из этих общих соображений, авторы изначально предполагали, что параллельное появление бескрылого фенотипа на пяти «небесных островах» Аризоны было связано с одними и теми же мутациями, изменившими ход развития крыльев у самок одинаковым образом. Это, однако, не подтвердилось.
Авторы изучили имагинальные крыловые диски (особые группы клеток личинки, из которых затем развиваются крылья) у будущих бескрылых самок с разных гор. Оказалось, что развитие крыловых зачатков у самок с северных и южных гор затормаживается на разных этапах. У будущих бескрылых самок с северных гор крыловые имагинальные диски на стадии взрослой личинки состоят из небольшого числа медленно делящихся клеток, тогда как у самок с южных гор на той же стадии крыловые диски заметно крупнее, а клетки в них делятся чаще.
Авторы также исследовали экспрессию шести ключевых генов, управляющих развитием крыльев. Эти гены входят в состав эволюционно консервативной генной сети, которая мало менялась на протяжении примерно 325 млн лет эволюции насекомых с полным превращением (рис. 2).
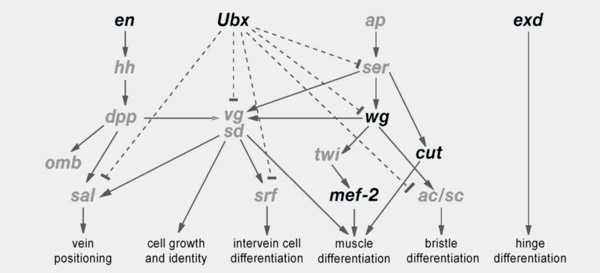
Рис. 2. Эволюционно консервативная (очень медленно меняющаяся в ходе эволюции) генная сеть, управляющая развитием крыльев у насекомых с полным превращением. Стрелочками показаны положительные воздействия (активация), тупичками — отрицательные (ингибирование). Жирным шрифтом выделены названия шести генов, экспрессия которых была изучена у самок Monomorium emersoni. Рисунок из обсуждаемой статьи в BMC Evolutionary Biology
Паттерн экспрессии этих генов в крыловых дисках (то есть пространственное распределение областей, где эти гены работают) сходен у крылатых самок и самцов разных муравьев (включая M. emersoni), дрозофилы и других насекомых. У бескрылых самок M. emersoni экспрессия этих генов в крыловых дисках, как и следовало ожидать, оказалась нарушенной. При этом у четырех генов, участвующих в пространственной разметке будущего крыла (ubx, en, cut, wg), характер нарушений оказался одинаковым у бескрылых самок с разных гор. Напротив, экспрессия гена mef-2, регулирующего развитие крыловой мускулатуры, изменилась по-разному у бескрылых самок южных и северных гор (хотя крыловая мускулатура в итоге не развивается ни у тех, ни у других). Наконец, экспрессия гена exd, регулирующего развитие шарнира основания крыла (hinge), изменилась одним способом на одной северной горе и одной южной, а на остальных горах она изменилась другим способом. Форма рудиментарных шарниров, имеющихся у бескрылых самок, чуть-чуть различается на разных горах в соответствии с характером экспрессии exd.
Таким образом, исследование показало, что быстрая параллельная эволюция важного нового признака (бескрылых самок), вызванная климатическими изменениями и фрагментацией местообитаний, была в основном закономерной и предсказуемой, но содержала также и элементы случайности. Закономерные (повторяющиеся) аспекты эволюционных изменений были связаны не только со сходными условиями среды на пяти «небесных островах» Аризоны, но и со сходным эволюционным потенциалом пяти популяций, который, в свою очередь, определялся общей генетической основой регуляции индивидуального развития. Элемент случайности, в свою очередь, был связан с историей популяций, которые за время, прошедшее после их разделения, могли накопить небольшие нейтральные различия в регуляторных генных сетях. Эти различия, возможно, поначалу вовсе не проявлялись в фенотипе, но впоследствии могли повлиять на то, какие именно мутации, ведущие к редукции крыльев у самок, «подвернутся под руку» естественному отбору в нужный момент.
Источник: Marie-Julie Favé, Robert A. Johnson, Stefan Cover, Stephan Handschuh, Brian D. Metscher, Gerd B. Müller, Shyamalika Gopalan and Ehab Abouheif. Past climate change on Sky Islands drives novelty in a core developmental gene network and its phenotype // BMC Evolutionary Biology. 2015. V. 15. P. 183.
См. также об эволюции на островах:
1) Эволюция на островах идет быстрее, «Элементы», 14.09.2006.
2) Видообразование на разных островах идет параллельными путями, «Элементы», 15.03.2007.
О соотношении случайного и закономерного в эволюции:
1) Анализ геномов калифорнийских палочников помог оценить соотношение случайности и закономерности в эволюции, «Элементы», 20.05.2014.
2) Цихлиды — живая модель независимой параллельной эволюции, «Элементы», 14.11.2012.
3) Параллельная эволюция эусоциальности у пчел: принцип общий, эволюционные маршруты — разные, «Элементы», 15.05.2015.
4) Параллельная эволюция изучена в эксперименте на бактериях, «Элементы», 01.02.2012.
5) Предсказуемый рост приспособленности достигается непредсказуемыми путями, «Элементы», 30.06.2014.
-
Решительно не понял, где здесь элемент случайности и почему этот якобы "элемент" - следствие разного пула нейтральных мутаций, накопленных популяциями за время изоляции. Во-первых, степень разности этого пула не легко, а очень легко оценить экспериментально, хотя бы допотопным, то бишь догеномным методом полиморфизма длины рестрикционных фрагментов. Во-вторых, достаточно взглянуть на приведенную схему, чтобы убедиться, что гены ubx, en, cut, wg работают в единой генной сети, а остальные - нет. В третьих, заглянув в допотопный учебник Рэффа и Кофмана из догеномных 80 гг. аж прошлого века можно узнать, что все эти гены работают в очень-очень-очень раннем эмбриогенезе, когда еще и имагиальных дисков крыла никаких нет. Из пункта 2 и 3 следует, что именно у этих генов пространство возможных - совместимых с жизнью - эволюционных изменений будет меньше, чем у остальных приведенных здесь генов. Вывод - никакого случайного элемента в том, что, характер нарушений именно у ubx, en, cut, wg оказался одинаковым у бескрылых самок с разных гор нет. Никакого случайного элемента в том, что mef-2 изменился всего двумя разными способами тоже нет - он работает в той же сети, но на самых поздних стадиях. Кроме того, он регулирует летную мускулатуру, а все, кто знаком с анатомией насекомых (учебник Бей-Биенко аж 60 гг. прошлого века) знает, что это - несколько специализированных тяжей, проходящих именно так, а не иначе. Поэтому сломать ее можно только ограниченным количеством способов, а избавиться от нее совсем нельзя - у полового поколения муравьев это еще и НЗ, горба у них нет, не верблюды, чай. Вот если бы mef-2 изменился не двумя, а 22 различными способами, это однозначно говорило бы и о элементе случайности и о его связи с разным пулом нейтральных мутаций. А так - получается именно так и должно быть, если никакой случайности и нейтральности нет (особенно если вспомнить, что все популяции принадлежат к одному виду). Складывается впечатление, что случайность и нейтральность приведены здесь для красного словца - чтобы хоть что-то из современной СТЭ прислонить к Вавилову. Но Вавилов в этой подпорке не нуждается. Наоборот, СТЭ без Вавилова скоро развалится как развалилась некогда единая популяция вышеуказанного муравья Monomorium emersoni в Аризоне. Я могу понять авторов, которым нужно протащить статью через высокоимпактную англоязычную рецензию, где ссылка на работу русского автора образца 1922 г. с непонятным словом homologous (по-английски это значит и гомологичный, и гомологический, вавиловский смысл для тех англоязычных, кто не знает греческого, совершенно непонятен). Это будет как красная тряпка для быка. Но я не понимаю Александра, который одновременно пишет хорошие книги, и повторяет авторскую тень, наведенную на плетень совершенно ясных, четких, красивых и с вавиловской точки зрения совершенно предсказуемых результатов.
PS Большой Привет кунинской "Логике случая" :-)-
Они таки выполнили оценку пула геномных различий в популяциях. Был использован метод Полиморфизма длины амплифицированных фрагментов, 4-мя парами праймеров, с получением на выходе 157 локусов (см. Материалы и методы в оригинале).
-
Итак, о Законе гомологических рядов Вы знаете не больше авторов статьи. Перейдем теперь к нейтральным мутациям. Прелесть нейтральных мутаций в том, что они фиксируются с той же скоростью, с какой и возникают. Поэтому их частота зависят от мутабильности и количества поколений. Ваше требование зависимости частоты нейтральных мутаций от количества изолированных популяций ("рассматривалось всего 5 изолированного эволюционирующих популяций, так что больше и неоткуда") - абсурдно. Очевидно Вы исходили из того, что мутабильность столь низка, что вероятнее всего в каждой из популяций после расщепления общей популяции зафиксировалось лишь одно нейтральное изменение гена mef-2. Но и это абсурд - авторы оценили мутабильность, как Вы сами и отметили. Именно поэтому я написал 22, (хотя мог бы написать и 3, и 6, и 8) - заведомо больше, чем по одной мутации на популяцию.
-
Нет, я не имела ввиду 5 мутаций, я говорю о 5 немного разных способах (на уровне экспрессии генов и онтогенеза) получения схожего фенотипа. Их в этом исследовании больше 5 быть не могло. А о возможном влиянии предшествующих нейтральных мутаций (кстати, необязательно затрагивающих те же самые гены) говорит, например, то, что один из генов развития крыла изменил характер экспрессии в противоположных направлениях, хотя с точки зрения банальной логики это вроде бы нелогично, ведь нужен одноправленный конечный результат. Конечно, в рамках этого исследования про нейтральные мутации — это только спекуляции (размышления то есть) но разве это запрещено. Главное, чтобы конкретные результаты и доказанные положения достаточно четко определялись от спекуляций. Мне показалось, что в данном случае это требование вполне соблюдено.
-
-
-


-
-
-
А по-моему, акценты расставлены очень даже верно. Видеть здесь только гомологический ряд Вавилова и ничего больше — просто какая-то упрямая односторонность. Параллелизм возникновения однотипного признака родственных организмов в сходном градиенте среды налицо, здесь все по Вавилову (согласна, что было бы приятно, если бы его упомянули) и это первый важный вывод, который авторы особо подчеркивают. И то, что изменения затрагивают одни и те же гены, участвующие в соответствующих стадиях развития. Но о полном детерминизме можно было бы говорить только если бы во всех пяти случаях развитие нового признака обеспечивалось одинаковым новым способом развития и одинаковыми новыми вариантами данных генов, а вот этого как раз таки и нет. Какая разница, обнаружилось ли 5 разных способов или 25 (собственно рассматривалось всего 5 изолированного эволюционирующих популяций, так что больше и неоткуда)? Может дадите свое, не легкомысленное объяснение?
-
1) "Параллелизм возникновения однотипного признака родственных организмов в сходном градиенте среды налицо, здесь все по Вавилову" - это как раз не по Вавилову, а по Дарвину. Это конвергенция, а не гомологический ряд. В законе гомологических рядов ни слова не сказано про общую среду.
2) "Но о полном детерминизме можно было бы говорить только если бы во всех пяти случаях развитие нового признака обеспечивалось одинаковым новым способом развития и одинаковыми новыми вариантами данных генов, а вот этого как раз таки и нет" - Гомологический - это сходство по принципу орнанизации, а гомологичный - по происхождению. Ваше требование полной идентичности, которого "как раз таки и нет" - это требование гомологии, а не гомологичности. Гомологический ряд в фенотипе возникает по Вавилову на базе общего сходства набора генов-гомологов, дополненного общим сходством физиологического биохимического и онтогенетического пути от гена до признака. Последнее и есть сходство генной сети, говоря современным языком. Общего сходства, а не идентичности! Вавилов подчеркивал, что сводить гомологический ряд к одному и тому же набору генов-гомологов - это ошибка. Которую Вы и совершили. Не то, что идентичности, но даже большого сходства генов-гомологов может и не быть, но сходства в организации того, как эти гены работают достаточно, чтобы у нескольких родственных таксонов появился один и тот же признак. Грубо говоря, у трех видов не работает ген А, но у одного вида мутация в промоторе гена А, у другого - в кодирующей части гена А, а у третьего роль гена А выполняет ген Б. Вот это и будет гомологический ряд: эквифинальность, когда один и тот же результат можно получить разными путями, лишь бы они затрагивали общий принцип организации.
-
-
-
Последние новости










Рис. 1. Пять «небесных островов» (sky islands) Аризоны и самки обитающих на них муравьев Monomorium emersoni: крылатая и бескрылая формы. Изображение из обсуждаемой статьи в BMC Evolutionary Biology