Бабочки монархи учились летать в Мексике

Бабочки монархи знамениты своими дальними путешествиями: эти легкие создания каждую осень летят из Канады в Мексику, преодолевая расстояние в три тысячи километров. Как появилась эта их удивительная особенность, очевидно гибельная для большинства особей? Предполагалось, что миграции бабочек возникли во время потепления климата, так что изначально оседлые бабочки постепенно распространялись к северу со своей экваториальной родины. Но генетический анализ, выполненный международной командой ученых, показал, что сценарий становления миграций был иным. Длительные миграции появились на основе коротких, которые были изначально характерны для популяций североамериканских субтропиков. Направленное изменение одного лишь гена способствовало укреплению мышц крыльев и, соответственно, позволяло выполнять всё более далекие перелеты. Этот ген четко выявляется при помощи известных методик изучения эволюции.
Миграции животных и история происхождения миграций до сих пор остаются интереснейшей загадкой для науки. Действительно, почему лососи упрямо возвращаются из моря в свою родную реку на нерест, проплывая для этого сотни километров, прыгая вверх по водопадам, преображаясь внешне и внутренне — и всё только затем, чтобы отнереститься в положенном месте и умереть? А угри пускаются в еще более долгое путешествие на 8000 км: стартуя из европейских рек, плывут с глубинными течениями к Саргассову морю. И, опять же, только затем, чтобы отложить икру и погибнуть среди бурых саргассов посреди Атлантики. Молодь угрей — лептоцефалы — влекомые Гольфстримом возвращаются в реки Европы, повторяя судьбу родителей. Мигрируют многие птицы, пролетая над океанами половину земного шара. А еще можно вспомнить миграции леммингов и саранчи, поразительные по своей массовости и расстояниям... И в каждом случае встает вопрос: почему животные пускаются в путь? Ведь часто это невыгодно с точки зрения выживания: по ходу долгого и нелегкого маршрута погибает значительная доля путешественников, до конечной точки добираются немногие.
Решать эту эволюционную загадку можно с помощью генетики: посмотреть, какие гены изменяются, чтобы обеспечить мигрантное поведение вида. Именно так и поступили генетики, взяв за основу «гены миграций» у бабочек монархов. Для выполнения этой работы собралась международная команда из нескольких научных лабораторий мира: Шанхайского института физиологии растений и экологии (Китай), Чикагского университета (США), Медицинской школы Массачусетского университета (Вустер, США), Стэнфордского университета (США), Хельсинкского университета (Финляндия), Университета Кордовы (Испания), Квинслендского университета (Брисбен, Австралия).
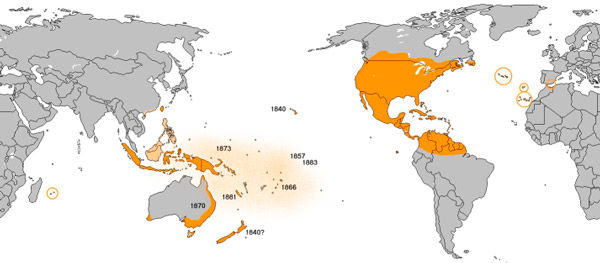
Ареал данаид монархов — Центральная и Южная Америка, Северная Америка до умеренных районов Канады, субтропические и тропические острова Тихого и Атлантического океанов, Южная Европа и Северная Африка. Наиболее дальние сезонные миграции свойственны североамериканским монархам. Карта с сайта en.wikipedia.org
Бабочки монархи Danaus plexippus представляют собой превосходный пример мигрантного вида. Эти нежные создания с легкими крылышками могут перемещаться во время сезонных миграций на 3000 км, переправляясь из умеренных районов Канады в Мексику. Там они зимуют и не размножаются. Зато весной, в марте, миллионы их собираются в стаи и устремляются обратно к северу; оставляя на своем пути яйца на развод следующих поколений, они погибают. Их дети и внуки продолжают неудержимое движение на север, также обзаведясь по пути потомством. До исходных северных территорий добирается уже третье или четвертое поколение монархов, отправившихся осенью в Мексику. Также монархи летят из Мексики в Южную Америку, известны трансатлантические перелеты монархов, мигрируют и популяции монархов Африки и Австралии. У Набокова найдем: «Бабочка стремится на юг, на зимовку, но разумеется гибнет, не долетев до тепла».
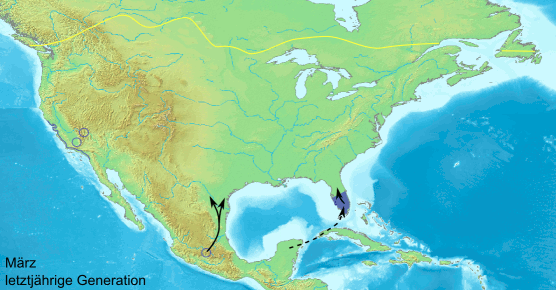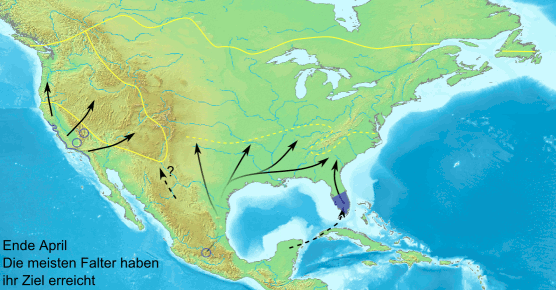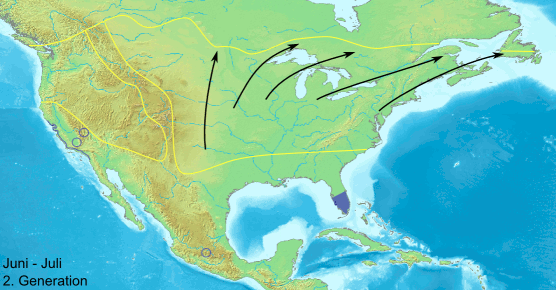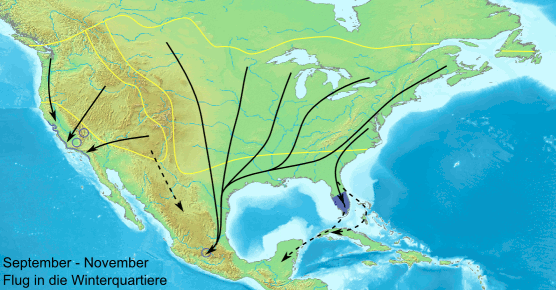
Сезонная миграция данаид монархов в Северной Америке устроена довольно сложно: бабочки с территорий восточнее Скалистых гор направляются зимовать в Мексику, а обитатели запада — в Калифорнию. Весной они начинают обратный путь, но до Скалистых гор долетают лишь внуки и правнуки осенних мигрантов. Анимация с сайта en.wikipedia.org
Как естественный отбор поддержал такую изумительную расточительность, воспитав в этих легких созданиях выносливость и целеустремленность? Ведь большинство видов этого рода (Danaus), равно как и многие популяции монархов, оседлые и никуда не мигрируют, обосновавшись в тропиках и экваториальной части материков. Этим популяциям незачем пускаться в путь, они могут спокойно размножаться, оставляя несколько поколений потомства ежегодно в одном и том же месте.
Миграция монархов — явление одновременно зрелищное и загадочное — вызывает немало споров среди ученых. Классическое объяснение предполагает, что этот вид появился в экваториальной зоне Центральной Америки. Основу его составляли оседлые популяции. При наступившем потеплении после конца ледникового периода и возрастании численности особей вид стал расширять свой ареал, переселяясь в том числе и в северные районы Америки. Однако там условия не подходили для зимовки, поэтому северные популяции были вынуждены выработать навыки длительных путешествий, преобразовав для этого свой жизненный цикл. Такая интерпретация смотрится логично. Но оказалось, что здравое логичное объяснение не подтверждается генетическими фактами. Дело обстояло как раз наоборот: мигрантные особи американских субтропиков дали начало оседлым популяциям тропиков и экватора, а также расселись по всему миру.
Чтобы выяснить происхождение миграций, ученые проанализировали около сотни геномов монархов и их ближайших родичей из разных популяций, мигрантных и оседлых. У мигрирующих популяций Мексики (мигрирующих хотя бы на короткие дистанции) генетическая вариабельность по однонуклеотидным заменам оказалась гораздо выше, чем во всех других частях света, а также выше, чем у оседлых соседей. Кроме того, у мексиканских популяций оказалась повышена доля гетерозиготных аллелей по многим генам, тогда как у популяций Центральной и Южной Америки, Европы, Африки и островов карибского бассейна и Тихого океана выше была доля гомозиготных генов. Доля гомозигот увеличивалась по мере увеличения удаленности популяций от Мексики. Это позволило предположить, что бабочки монархи начали свой эволюционный путь именно из Мексики, от популяций, совершающих короткие сезонные миграции. Они расширяли свой ареал вместе с распространением основного кормового растения — ядовитого ластовника Asclepias.
Мексиканские бабочки монархи стали расселяться из Мексики в Центральную Америку, оттуда в Южную Америку, попутно обосновавшись на островах Карибского бассейна. Другая волна расселения отправила бабочек через Атлантику в район современной Португалии, а оттуда условия позволили бабочкам перелететь в Южную Европу и Северную Африку. Считалось, что монархи появились в Европе лишь в начале XIX века, но, судя по генетическим маркерам, этот «исход» бабочек из Мексики в Европу через острова начался существенно раньше. Третья часть переселенцев освоила тихоокеанские маршруты, объявившись на островах Тихого океана, в Австралии и Новой Зеландии. По мере удаления от центра расселения — от Мексики — популяции оказывались в неблагоприятных, непривычных условиях, их численность резко снижалась, и в результате они теряли внушительную часть исходного генетического разнообразия. Иными словами, исходно мигрантные популяции эволюционировали в сторону увеличения дальности перелетов и использовали эту свою способность для расселения по всему свету. Также они дали начало и оседлым популяциям.
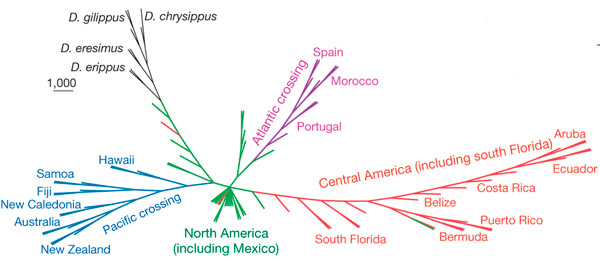
Филогенетическое дерево, построенное на основе анализа однонуклеотидных замен 89 популяций монархов и их ближайших родственников. Оно отражает пути расселения бабочек из субтропиков северной Америки в другие части света. Рисунок из обсуждаемой статьи в Nature
Что же помогло исходным мигрантам развить свою страсть к путешествиям? Как выяснилось, различия оседлых и мигрантных особей обнаруживаются в нескольких генах. Но исследователи отметили один из них с наиболее четкими следами движущего отбора. Так, участок последовательности вокруг нового, «мигрантного», варианта характеризуется сниженной генетической вариабельностью — это показатель недавней дивергенции. Это ген альфа-субъединицы коллагена IV (Collagen alpha-3(IV) chain). Этот белок определяет структуру мышц крыльев, вероятно укрепляя их. Также обнаружилась и высокая эффективность работы этих мышц: у мигрантов мышцы, связанные с полетом, потребляют гораздо меньше энергии. Это позволяет бабочкам лететь без отдыха до тысячи километров. У оседлых особей уровень метаболизма в мышцах крыльев гораздо выше, чем у мигрантов. По-видимому, «мигрантный» уровень метаболизма в обычной жизни вреден, поэтому все оседлые популяции переключились на более высокий уровень метаболизма. При этом остальные мышцы тела и конечностей работают у мигрантов и оседлых особей одинаково.
Этот пример показателен в нескольких аспектах. Во-первых, философская сторона дела: в природе не всегда верно то, что на первый взгляд кажется логичнее всего. Во-вторых, эволюционный урок: длительные миграции монархов появились не на пустом месте, а благодаря существующему уже заделу: способности к коротким сезонным миграциям. Стимулом для становления длительных миграций было, по-видимому, распространение основного кормового растения этого вида. Для вырабатывания навыков длительных полетов оказалось достаточным изменение небольшого числа генов, регулирующих структуру мышц крыльев и эффективность их метаболизма. Эти гены четко выявляются при помощи известных методик изучения генетической эволюции.
Источник: Shuai Zhan, Wei Zhang, Kristjan Niitepõld, Jeremy Hsu, Juan Fernández Haeger, Myron P. Zalucki, Sonia Altizer, Jacobus C. de Roode, Steven M. Reppert & Marcus R. Kronforst. The genetics of monarch butterfly migration and warning colouration // Nature. 2014. DOI:10.1038/nature13812.
Елена Наймарк
-
По поводу старого объяснения, мне кажется оно не такое логичное, так как короткие миграции не перенесут бабочек в лучшие климатические условия другого пояса, и наверно не закрепятся отбором поэтому. Мне кажется логичнее что в начале были местные миграции не связанные с глобальными климатическими зонами, а из-за локальных отличий местного климата, наличия еды или воды.
-
Странно, что не рассмотрена "паразитически-хищническая" теория, которая, собственно, и говорит, что размножаться лучше, где хищников и паразитов поменьше, но отжираться потом и "коротать дни" между стадиями размножения - где жратвы побольше. Первое - на севере, второе - на юге, следовательно, стимул к удлинению миграций. Понятно, к бабочкам, для которых весь маршрут миграции имеет временную длительность в несколько поколений, это применить, на первый взгляд, трудно, но и тут - размножение именно на северной части маршрута на любом из участков перелета. А несколько таких участков - это, простите, попросту выгодно для максимального распространения по широчайшему ареалу обитания, что несомненный плюс. Видимо, именно этот, последний, фактор и приводит к наличию "многостадийности" миграции в данном варианте.
-
Вот именно! Выгодно - это ключевое понятие для природы. Автор пишет, что "философская сторона дела: в природе не всегда верно то, что на первый взгляд кажется логичнее всего". Логичнее всего - с чьей позиции? Если с позиции выгоды, то всё выжившее - верно, каким бы сложным и многоступенчатым оно не казалось.
-
Тоже неверно точнее - неполно. Выгода - это механистичное, прямое толкование. На самом деле гораздо более корректной будет формулировка "вероятность существования отдалённого потомства", причём отдалённой следует считать,например, вечность :-)
А выгоды или смысла - в природе не существует.
Разверну: сине-зелёные водоросли существуют спокон веков, в чём же тогда смысл эволюции? А в том, что растёт вероятность сохранения генов, носимых данным конкретным экземпляром. Цианобактерии как были, так и есть - но вероятность того, что гены именно данной одной конкретной клетки будут существовать хотя бы через десять секунд - невелика.
А у, скажем, ворона, она практически 100%. И есть даже изрядная вероятность того, что его гены будут носимы и через 100 лет, и 200.
И дело не в целях или смысле (что одно и то же и чего в природе в принципе не существует). Дело просто в том, что те, кто не оставлял хоть какое (вовсе не обязательно многочисленное) потомство в течение всех тысяч и даже миллионов лет (без каких-либо перерывов :-) - исчезли. Собственно, это синоним фразы "для конкретного живого существа вероятность существования отдалённого потомства в ходе эволюции растёт", и это же можно считать формулировкой биологической эволюции.
Вернувшись к статье - стоит оценивать смысл дальних миграций именно в смысле вероятности оставить потомство, а не просто вероятность погибнуть.-
Разверну: сине-зелёные водоросли существуют спокон веков, в чём же тогда смысл эволюции? А в том, что растёт вероятность сохранения генов, носимых данным конкретным экземпляром - увы, Ваше объяснение не проходит, как раз гены меняются гораздо быстрее морфологии. И аналогия с вороном не проходит - ворон может упасть и разбиться, умереть с голоду, подавиться костью, намокнуть и умереть от банальной простуды. Для цианобактерии все эти причины ПРОСТО НЕ СУЩЕСТВУЮТ. Получается, что защищаясь от одной причины смерти ворон одновременно подставляется под сотни других. Это все равно, что защищаться от снайпера, ползя с гранатой под гусеницу вражеского танка.
-
Да пусть себе гены меняются, это-то при чём? Вопрос в вероятности существования именно этого гена в будущем. Если не существует - всё, нечего обсуждать. Причём ген "не знает" ничего о существовании своих копий в других особях. (он вообще ничего не знает :-), и в данный момент существуют только те, что существуют в течение миллионов лет _непрерывно_.
Считается вероятность банально просто: количество цианобактерий, как и воронов, можно считать примерно постоянным на интервале в год.
Ворон размножается раз в год, пара откладывает максимум 7 яиц. Если бы все всегда выживали, что воронов бы стало в четыре с половиной раза больше (эта пара плюс 7 - то есть 9 особей вместо исходных двух). А их на самом деле столько же. Значит, вероятность выживания 1/4,5=0.22
А цианобактерии делятся в среднем раз в сутки. То есть через год их должно быть 2^365 (два в степени триста шестьдесят пять, в то время как число атомов во вселенной примерно 2^250). Вероятность существования данной конкретной бактерии, соответственно, единица, делённая на это невообразимое число, то есть невообразимо близка к нулю. Хотя и не точно равна, потому-то цианобактерии до сих пор существуют :-)
В этом-то и "смысл" развития живых существ. Тех генов (и их носителей), вероятность существования которых во времени выше, становится больше и больше.
-
-
-
-
-
1) Это было бы замечательно, если бы бабочка смогла оценить "где паразитов и хищников поменьше". В реальности же монархи летят не просто из Канады в Мексику, а из Канады на конкретные деревья Мексики. Если же это дерево будет отсутствовать (сгорело, буря свалила) - бабочки очень неохотно начнут искать другое поблизости. Так что хищнику - только открывай рот. Но монарху вообще наплевать на хищника - бабочка ядовита. Размножение же на северном участке объясняется не отсутствием хищников, а наличием кормовых растений. Именно отсутствие корма для гусениц объясняет, почему монархи до сих пор не закрепились в Европе. Летают они через Атлантику довольно часто.
2)"Это, простите, попросту выгодно для максимального распространения по широчайшему ареалу обитания, что несомненный плюс" - если плюс столь несомненен, оседлые монарх просто не возникли бы.
3) Референт не заметил логического круга. Он пишет, что ничего нового не возникает само по себе, длинные миграции возникли на базе коротких. Тогда прошу объяснить возникновение коротких миграций, если существуют и процветают оседлые монархи. Короткая миграция - это как раз неустойчивое состояние между двумя крайними - оседлостью и дальней миграцией. Референт, конечно, может прибегнуть к спасительной "широкой норме реакции", но тогда ему придется объяснять возникновение этой нормы, если опять-таки оседлые популяции есть и процветают.


Последние новости






Миллионы бабочек монархов отправляются осенью из района Скалистых гор в Мексику, где проводят зиму. Они плотными гроздями облепляют хвойные или эвкалиптовые деревья и впадают в спячку. Фото с сайта webdiscover.ru