Программы работы Hox-генов у личинок и взрослых особей кольчатого червя принципиально отличаются

В современной биологии очень распространены исследования генов семейства Hox, которые являются важными регуляторами индивидуального развития животных. Лучше всего функции этих генов изучены у мухи-дрозофилы, на примере которой они и были открыты. Однако несмотря на то, что Hox-гены есть у подавляющего большинства современных животных, работать они могут по-разному. Если у личинки кольчатого червя нереиса механизм действия Hox-генов вполне похож на тот, что известен по дрозофиле, то у взрослого нереиса он оказался совершенно другим. Какой из этих вариантов эволюционно первичен — пока неизвестно.
Гены семейства Hox (очень часто их называют просто Hox-генами) — довольно большая группа генов, влияющих на индивидуальное развитие животных. Эти гены контролируют развитие разных частей тела: у насекомых, например, мутации Hox-генов могут вызывать появление добавочной пары крыльев, превращение усиков в лапки или изменение числа конечностей. Продуктами Hox-генов являются регуляторные белки, влияющие на работу каких-то других генов (такие белки называются факторами транскрипции).
Каждому Hox-гену свойственна своя область экспрессии, то есть часть организма, в которой активен этот ген и синтезируется его продукт. У сегментированного животного это может быть определенный сегмент или группа сегментов (рис. 1). Разные сегменты часто характеризуются разным набором продуктов Hox-генов; в некотором смысле, это и есть причина различий между сегментами. Например, у насекомых в 1-м, 2-м и 3-м грудных сегментах набор продуктов Hox-генов разный, и известно, что именно этими генами определяется их дифференцировка.

Рис. 1. Экспрессия Hox-генов в теле сегментированного животного. В первом сегменте экспрессируется только ген Hox2, во втором — Hox2 и Hox3, в третьем — Hox2, Hox3 и Hox4, в четвертом — Hox2, Hox3, Hox4 и Hox5. В переднем несегментированном отделе (белый цвет) экспрессии этих генов нет вообще. Прямоугольники справа обозначают протяженность областей экспрессии. Схема составлена по статье Shigeru Kuratani, 2005 в Journal of Anatomy
Количество Hox-генов в геноме бывает различным, но чаще всего их где-то около десятка: например, у мухи-дрозофилы — 8, у ланцетника — 14. Причем даже у таких разных животных большинство из этих генов являются общими. Hox-гены очень эволюционно консервативны.
В современной биологии исследования Hox-генов чрезвычайно популярны. Но львиная доля этих исследований касается всего двух групп животных: членистоногих и позвоночных. К членистоногим относится муха-дрозофила — самый заслуженный объект генетиков; первые открытия, связанные с Hox-генами, были сделаны именно на ней, и неудивительно, что ее изучение активно продолжается. Ну а позвоночные — это мы сами и наши родственники, уже поэтому не избежавшие внимания ученых. Кроме того, у позвоночных число Hox-генов необычно увеличено по сравнению с любыми другими животными (например, у человека их 39), и это тоже стимулирует интерес генетиков к данной группе. Наконец, и у членистоногих, и у позвоночных в теле очень хорошо выражена сегментация, а как раз у сегментированных животных эффекты Hox-генов проявляются особенно наглядно.
Между тем есть как минимум еще одна большая группа сегментированных животных: кольчатые черви. Сегментация кольчатых червей — самая «классическая», она выражена очень ярко и затрагивает, пожалуй, максимальное число систем организма. У таких животных действие Hox-генов, которые в первую очередь регулируют именно сегментацию, должно проявляться вовсю. Но известно об этом до сих пор маловато.
Группа исследователей из Лаборатории экспериментальной эмбриологии кафедры эмбриологии Санкт-Петербургского государственного университета изучила работу Hox-генов в развитии морского кольчатого червя Alitta virens. Этот червь известен также под старым названием Nereis virens, а по-русски его обычно называют просто нереисом.
Нереис был выбран в качестве объекта по следующим причинам:
1. Он относится к классу многощетинковых червей, который находится в основании эволюционного древа кольчатых, а потому, вероятно, сохранил черты общего предка этого типа.
2. У него есть хорошо выраженная личиночная стадия, так что можно сравнить работу Hox-генов у личинки и у взрослого червя.
3. Его многочисленные сегменты очень похожи один на другой — тело не подразделено на отделы (в отличие от тел как членистоногих, так и позвоночных). Надо сказать, что есть кольчатые черви, у которых сегменты в той или иной степени дифференцированы, но именно у нереиса этого почти совсем нет. Интерес исследователей как раз и был в том, чтобы выяснить, как работают Hox-гены в таком организме, где все сегменты одинаковы.

Рис. 2. Личинка-нектохета (слева) и взрослый нереис (справа). lh — личиночная голова, ah — голова взрослого червя, GZ — задняя зона роста, в которой возникают новые сегменты, cI — усики (цирры) околоротового отдела, cII — усики (цирры) первого личиночного сегмента, звездочка — рот. Серый цвет обозначает части тела, которые есть у личинки, белый — части, возникающие в постларвальном (послеличиночном) развитии. У взрослого червя первый личиночный сегмент прирастает к переднему несегментированному отделу, входя в состав головы. Рисунок из обсуждаемой статьи в EvoDevo
Для выявления областей экспрессии Hox-генов у червя был применен широко распространенный в наше время метод гибридизации in situ на тотальном препарате. Суть этого метода следующая. Непосредственным продуктом гена является информационная РНК (иРНК), с помощью которой потом синтезируется белок. Зная нуклеотидную последовательность иРНК данного гена, можно искусственно синтезировать ДНК-зонд — цепочку нуклеиновой кислоты, которая будет специфически (комплементарно) связываться с этой иРНК, и только с ней. Если химически присоединить к этому зонду светящуюся метку, то, обработав нужными компонентами ткань, орган или даже целое тело интересующего нас животного (в последнем случае как раз и говорят о «тотальном препарате»), мы можем увидеть там светящуюся зону. Это и будет область экспрессии гена.
Первым объектом изучения питерских исследователей стала поздняя личинка нереиса — нектохета (рис. 2). Ее тело состоит из переднего и заднего несегментированных отделов, между которыми находятся три личиночных сегмента. В несегментированных отделах экспрессии Hox-генов почти нет. В сегментах же она выражена очень четко. Например, в первом личиночном сегменте экспрессируется ген Hox1, во втором к нему присоединяется Hox4, а в третьем еще и Hox5 (рис. 3, верхняя диаграмма). Таким образом, из трех перечисленных генов в первом личиночном сегменте экспрессируется один, во втором два, а в третьем все три. Это очень напоминает «классическую», изученную на насекомых и позвоночных модель, где каждый сегмент характеризуется собственным набором продуктов Hox-генов, и чем дальше по телу назад, тем этих продуктов больше (рис. 1).
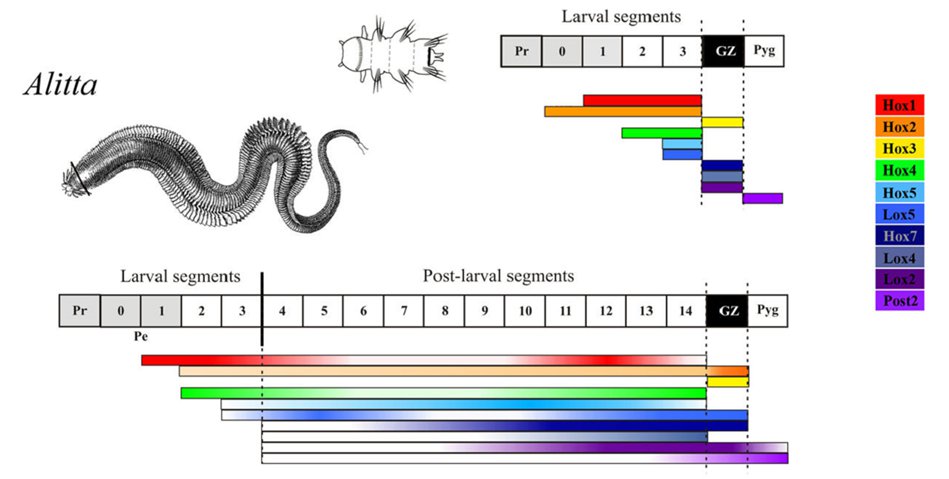
Рис. 3. Экспрессия Hox-генов у нектохеты нереиса (вверху) и у взрослого нереиса (внизу). Pr — простомиум, Pyg — пигидиум, Pe — перистомиум, GZ — зона роста, цифры — номера сегментов. Простомиум, пигидиум и нулевой сегмент (относящийся к перистомиуму) в тексте не обсуждаются. Столбики на диаграммах обозначают зоны экспрессии отдельных генов, как на рис. 1. Интенсивность цвета соответствует силе экспрессии данного гена в данной области. Рисунок из обсуждаемой статьи в EvoDevo
У взрослого червя картина резко меняется. Оказывается, все его сегменты, образовавшиеся после личиночной стадии (они называются постларвальными), практически вообще не отличаются друг от друга по набору продуктов Hox-генов (рис. 3, нижняя диаграмма). Только в первых трех сегментах — ларвальных — эти отличия сохраняются. Но ларвальные сегменты составляют очень маленькую часть тела взрослого червя: новые, постларвальные, сегменты образуются у нереиса всю жизнь, так что их вполне может быть около 200.
Сравнивая личинку нереиса со взрослым червем, мы увидим следующее. По окончании личиночной стадии зоны экспрессии «передних» Hox-генов (например, уже упомянутых генов Hox1, Hox4 и Hox5) просто распространяются назад, охватывая все постларвальные сегменты. Кроме того, у личинки есть гены, которые экспрессируются только в задней зоне роста (она находится позади последнего сегмента). Это, например, ген Hox7. У взрослого червя зона его экспрессии распространяется вперед и тоже охватывает все постларвальные сегменты (или, во всяком случае, почти все).
Есть ли вообще между постларвальными сегментами нереиса различия, связанные с экспрессией Hox-генов? Да, есть. Но они заключаются не в наборе продуктов этих генов, а только в их концентрациях. Продукт каждого из генов, перечисленных выше, есть во всех постларвальных сегментах, но где-то его концентрация может быть больше, а где-то меньше. Например, продукт гена Hox4 дает два пика концентрации, впереди и сзади (рис. 4).
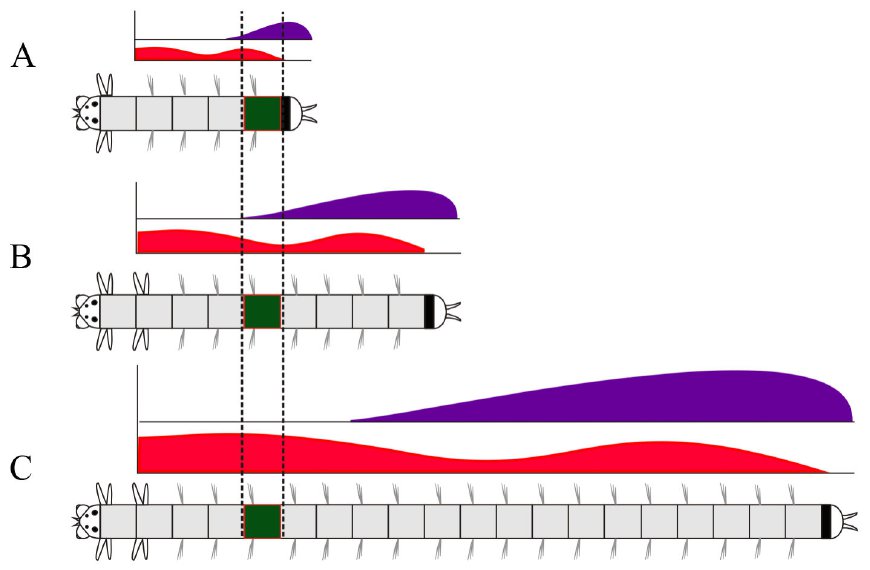
Рис. 4. Изменения экспрессии двух Hox-генов в одном постларвальном сегменте за время роста червя. Произвольно выбранный сегмент выделен темно-зеленым цветом. В постларвальном развитии кольчатых червей новые сегменты возникают только в задней зоне роста (черная полоска позади последнего сегмента). Красным цветом показана концентрация продукта гена Hox4, фиолетовым — продукта гена Post2. Когда сегмент только что образовался, он находится у самой зоны роста. В это время уровень экспрессии и Hox4, и Post2 в нем высок (A). Однако в зоне роста продолжают возникать новые сегменты, так что «наш» сегмент оказывается в середине тела червя (B). В этот момент в нем низкая экспрессия как Hox4, так и Post2. И наконец, когда новых сегментов возникло уже столько, что «наш» сегмент сместился в переднюю часть червя (C), уровень экспрессии Hox4 в нем опять повышается. Рисунок из обсуждаемой статьи в EvoDevo
Анализируя эти данные, петербургские эмбриологи говорят о существовании у животных двух независимых морфогенетических программ — «ларвальной» и «постларвальной». Если в «ларвальной программе» различия между сегментами устроены по принципу «всё или ничего» (или экспрессия данного гена в данном сегменте есть, или нет), то в «постларвальной программе» они являются чисто количественными. Ситуация, с которой традиционно имели дело исследователи Hox-генов у членистоногих или у позвоночных, соответствует «ларвальной программе» (рис. 1). Но у нереиса «постларвальная программа» настолько явно доминирует, что изучать придется именно ее.
Какая морфогенетическая программа эволюционно древнее — «ларвальная» или «постларвальная»? На этот вопрос ответа пока нет. Но замечательно уже то, что теперь он четко поставлен.
«Ларвальная» и «постларвальная» программы — это принципиально разные способы позиционной разметки тела (см. Как клетки понимают, что одни должны стать волосами, другие костями, третьи мозгами и т. п.? И из какого центра им подаются команды? «Элементы», ответ А. Маркова на детский вопрос). И дело здесь не только в том, что в «постларвальной программе» не действует принцип «всё или ничего». Есть еще одна важная закономерность. В «ларвальной программе» каждый сегмент — например, 1-й или 3-й — характеризуется уникальным для него сочетанием продуктов Hox-генов. В «постларвальной программе» уникальные сочетания концентраций этих продуктов тоже присутствуют, но они зависят не от номера сегмента, а от расстояния данной точки тела животного от переднего и заднего концов. А вот в каждом отдельном сегменте эти концентрации как раз меняются по мере роста (рис. 4, A–C).
Петербургские авторы высказывают гипотезу, что «постларвальная программа» обеспечивает животному высокую способность к регенерации утраченных частей тела. Если какие-то сегменты потеряны, соотношение концентраций продуктов Hox-генов всё равно сохраняется (оно от наличия или отсутствия отдельных сегментов не зависит), и в результате делящиеся в области раны клетки «знают», сколько и каких сегментов они должны образовать. И действительно, регенерация у кольчатых червей прекрасно развита.
Животные, у которых исключительно хорошо развита регенерация, есть и среди позвоночных. Это хвостатые амфибии. Еще в 2003 году группа французских авторов показала, что у типичного представителя хвостатых амфибий, испанского ребристого тритона Pleurodeles waltl, сохраняется во взрослом состоянии высокая активность нескольких генов семейства Hox в спинном мозге. Эта особенность уже сама по себе не совсем заурядна, обычно Hox-гены активны преимущественно у зародышей. При этом в спинном мозге тритона зоны экспрессии Hox-генов очень широко перекрываются, а внутри этих зон имеются явные пики концентраций, для продукта каждого гена свои (см.: S. Nicolas et al., 2003. The spatial restrictions of 5'HoxC genes expression are maintained in adult newt spinal cord). Такая картина чрезвычайно напоминает «постларвальную программу», описанную теперь для нереиса. Французские авторы сразу предположили, что такой способ работы Hox-генов связан с высокой способностью животных к регенерации. Похоже, исследования кольчатых червей подтверждают эту идею.
Источник: Nadezhda I. Bakalenko, Elena L. Novikova, Alexander Y. Nesterenko and Milana A. Kulakova. Hox gene expression during postlarval development of the polychaete Alitta (Nereis) virens // EvoDevo. 2013. V. 4. P. 13. Статья в открытом доступе.
Сергей Ястребов
-
Хороший обзор автора сообщения, отличные результаты авторского коллектива статьи. Наблюдаю, как в последние 3-4 года питерские ученые реанимируют концепцию первичной гетерономности П.П.Иванова - непринятую за рубежом и постепенно забываемую у нас. Очень интересно. Но вот, что любопытно:
>Петербургские авторы высказывают гипотезу, что «постларвальная программа» обеспечивает животному высокую способность к регенерации утраченных частей тела.
Судя по всему, такая программа первична для аннелид - иначе трудно понять, почему у наиболее высокоразвитой аннелидной группы, пиявок, ее нет вообще. Но, если это так, то тогда мы в праве ожидать ее обнаружение и у членистоногих (тем более, что и у них есть первичная гетерономность). Есть ли данные о том, что у членистоногих (прежде всего, у ракообразных с науплиальной личинкой) экспрессия Hox в первых 3-5 сегментов отличается от остальных? Если "нет", то это может быть доказательством в "копилку" того, что сегментация у артропод и аннелид возникла независимо, а то, что авторы описали для нереиса - специфика аннелидной ветви.-
Насколько я понимаю, разница между "ларвальной" и "постларвальной" программами (в понимании Кулаковой с коллегами) возникает естественным образом у любого животного, у которого число сегментов значительно превосходит число Hox-генов. Там есть хитрости с разными ступенями экспрессии, но все равно на длинное тело их не хватит. Про ракообразных пока ничего не знаю, хотя опять же подозреваю, что это зависит просто-напросто от длины тела. Мне, в свою очередь, стало очень интересно, как с этими делами у многоножек. По ним работы точно есть, постараюсь добраться.
Что касается независимого происхождения сегментации, то вот, например:
http://onlinelibrary.wiley.com/doi/10.1002/bies.200900130/abstract
В последнее время гипотезы независимого рекрутирования одних и тех же генов для аналогичных функций постепенно становятся популярны.
-
-
Вот что еще интересно было бы - это найти животное, у которого _только_ постларвальная программа и ларвальной нет вообще. Но боюсь, что это морфогенетически запрещено. Надо же как-то размечать головной мозг.


Что интересно, пока мне не попалось ни одной работы, где обсуждалась бы работа Hox-генов у взрослых членистоногих. Хотя, что-то такое наверняка есть, поскольку у ракообразных бывают гетероморфозы - некорректная регенерация при которой вместо одной структуры отрастает другая, то есть происходит гомеозисная трансформация.
Наша полихета в отличие от многоножки растёт всю жизнь и постларвальная работа Hox-генов нужна ей для контроля над ростом и регенерацией. Ей нужно контролировать позицию многочисленных единообразных метамеров чтобы скоординировать их рост, дифференцировку и процесс формирования новых сегментов. Примечательно, что позиционная информация в стволовых клетках взрослых млекопитающих находится под контролем всё тех же Hox-ов. Такой вот привет из Кембрия. Да, а животное, которое пользуется только постларвальной программой есть - это планария. Сегментов у неё нет, а градиенты Hox-генов есть. И, по крайней мере один Hox-ген планарии участвует в регенерации ( http://onlinelibrary.wiley.com/doi/10.1046/j.1440-169X.2001.
Прошу простить за многословие и некоторую сумбурность. Очень приятно, что нашу работу заметили. Сергей, большое спасибо за внятный и умный обзор. Делать научно-популярные очерки чрезвычайно сложно. Я бы сама не взялась. Честно.
-
О, я тоже рад, что Вам понравилось.
По пунктам:
1. Про планарию действительно интересно. А у моллюсков работа Hox-генов ближе к ларвальной или к постларвальной модели? По теории первичной гетерономности, тело моллюсков - целиком ларвальное, но это мало что значит, т.к. мы до сих пор не знаем, какое состояние первично. Как раз тут очень нужны данные по генетике развития.
1а. А вообще, я мечтаю узнать, как работают Hox-гены у приапулид. Известно, что их очень трудно разводить, но кто первый сделает такую работу - сорвет банк. Самые базальные Ecdysozoa, безоговорочно кембрийская группа.
2. Да, у многоножек конечное число сегментов фиксировано, но у них довольно часто бывает анаморфное развитие, когда оно (число) увеличивается уже на взрослой стадии постепенно, пока не дойдет до финального. А еще у них бывают всякие любопытные усложнения, как, например, у диплопод. На эту тему есть интересные работы Минелли, но он сам совершенно не генетик.
3. Очень интересна судьба теории П.П. Иванова. Насколько я понимаю, эта теория была почти полностью забыта после работ по онихофорам, у которых ларвальных сегментов просто нет. И то, что вы возвращаете ее в научный оборот, замечательно.-
У изученных моллюсков программа скорее ларвальная - колинеарная экспрессия Hox-генов в нервной системе. А кроме того много коопций - Hox-гены участвуют в эмбриогенезе разных "моллюско-специфичных" структур, таких как раковинная железа или венчик щупалец.
Про приапулид действительно очень интересно. Интересно сколько у них Hox-генов и какие это гены.
-
Последние новости









Фотография червя Alitta (Nereis) virens с флуоресцентными метками, показывающими область экспрессии одного из Hox-генов (в данном случае гена Lox4). A — личинка-нектохета, B–E — молодой червь после метаморфоза. Рисунок из обсуждаемой статьи в EvoDevo