Хромосомные перестройки не мешают бурозубкам свободно скрещиваться

У многих организмов хромосомные перестройки вызывают генетическую дифференциацию, которая впоследствии приводит к видообразованию. Поэтому, как правило, число хромосом может сильно различаться у близкородственных видов, но мало варьирует внутри вида. Однако встречаются и исключения, к которым, в частности, относится обыкновенная бурозубка. У этого зверька описано более 70 хромосомных рас. Швейцарские ученые с соавторами из разных стран исследовали пять гибридных зон между хромосомными расами этого вида и показали, что зверьки из разных рас легко скрещиваются между собой. Примечательно, что различие по числу хромосом не препятствует скрещиванию разных рас, но является существенным барьером при скрещивании разных видов бурозубок.
Роль хромосомных перестроек в видообразовании активно обсуждается в литературе в последние десятилетия. Каким образом слияние или разрыв хромосом могут способствовать видообразованию? Это может происходить по двум причинам. Во-первых, гибриды, получившие разное число хромосом от отца и матери, могут оказаться нежизнеспособны или стерильны, так как в ходе мейоза хромосомы не находят себе пары. Во-вторых, может нарушаться рекомбинация (см. Genetic recombination) вблизи участков хромосом, где произошли перестройки; в результате сильно снижается обмен генами в этих областях хромосом, что также может приводить к генетической дифференциации, а в дальнейшем и к видообразованию. Видимо, в разных случаях либо первая, либо вторая причины являются первостепенными, однако до сих пор накоплено мало экспериментальных подтверждений.
Близкородственные виды из группы обыкновенной бурозубки, или Sorex araneus, являются прекрасным объектом для изучения влияния хромосомных перестроек на обмен генами. У этих мелких млекопитающих потрясающая вариабельность кариотипов, а у S. araneus описано до 70 хромосомных рас. Любопытно, что эта изменчивость достигается в основном за счет Робертсоновских транслокаций (см. Robertsonian translocation), когда длинные плечи негомологичных хромосом объединяются в одну хромосому, а короткие плечи теряются.
Для этой группы бурозубок описано несколько гибридных зон между различными расами (рис. 1). Гибриды различаются по степени сложности кариотипов, в зависимости от того, насколько сильно различаются родительские расы. В одних случаях кариотип гибридов состоит только из тривалентов (когда две хромосомы из одного набора спариваются с одной хромосомой из другого); такие гибриды называются «простыми гетерозиготами». В других случаях у гибридов могут присутствовать длинные цепочки из многих (до 11) слившихся хромосом; такие гибриды называются «сложными гетерозиготами».

Можно предположить, что поток генов в разных гибридных зонах S. araneus варьирует в зависимости от степени различий родительских кариотипов. Теоретически, он должен быть тем ниже, чем сложнее кариотип гетерозигот. Кроме того, поток генов между хромосомами с перестройками должен быть ниже, чем между обычными хромосомами. Эти две задачи были исследованы большим коллективом авторов из университетов Швейцарии, Канады, Польши, России (Института цитологии и генетики СО РАН, кафедры цитологии и генетики Новосибирского государственного университета и Института проблем экологии и эволюции им. А. Н. Северцова РАН), Великобритании, США и Чехии.
Для решения этих задач авторы исследовали 876 зверьков из пяти гибридных зон (рис. 1). В качестве генетических маркеров были выбраны 16 микросателлитов, расположенных как на обычных хромосомах, так и на хромосомах с перестройками. Для анализа генетической структуры популяций использовали так называемую F-статистику, или индекс фиксации. В популяционной генетике F-статистика описывает уровень гетерозиготности в популяции, или, точнее, степень снижения гетерозигот в популяции по сравнению с равновесием Харди—Вайнберга. Напомним, что, согласно закону Харди—Вайнберга, равновесие между частотами гомозигот и гетерозигот может быть в идеальной популяции, в которой не действует отбор и которая состоит из бесконечного числа свободно скрещивающихся особей. Индекс фиксации варьирует от 0 до 1, причем значение 0 говорит о генетической идентичности, а значение 1 говорит о том, что две популяции являются разными видами.
Что же дало проведенное исследование? Оказалось, что, независимо от уровня сложности гибридного кариотипа, во всех зонах были получены низкие значения индекса фиксации (рис. 2). Кроме того, индекс фиксации достоверно не различался между хромосомами с перестройками и обычными хромосомами. Таким образом, ни уровень сложности кариотипа, ни тип хромосом не влияли на генетическую дифференциацию между расами S. аraneus.
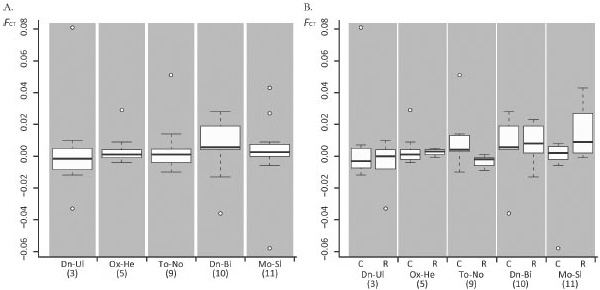
Эти результаты отличаются от ранее полученных данных по бурозубкам разных видов, а также по другим таксонам (например, дрозофилам, мышам или подсолнухам). Почему же различие по числу хромосом не препятствует скрещиванию разных рас S. аraneus, но является существенным барьером при гибридизации разных видов бурозубок — например, между S. araneus и S. antinorii в Альпах (Basset et al., 2006)? Авторы не находят убедительного ответа на этот вопрос.
В нескольких исследованиях было показано, что у гетерозигот в группе S. аraneus сохраняется высокая фертильность, но причина этого не ясна. Популяционный анализ по различным генетическим маркерам также не раз показывал очень низкий уровень генетической дифференциации между расами S. аraneus. Этот факт, по мнению авторов, объясняется тем, что кариотипы бурозубок в этой группе эволюционировали совсем недавно, примерно между 15 000 и 7000 годами до настоящего времени. Авторы считают, что в дальнейшем при проведении генетического анализа желательно знать точную локализацию микросателлитов на хромосомах, так как частота рекомбинаций может сильно варьировать в зависимости от позиции маркера.
Источник: A. Horn, P. Basset, G. Yannic, A. Banaszek, P. M. Borodin, et. al. Chromosomal rearrangements do not seem to affect the gene flow in hybrid zones between karyotypic races of the common shrew (Sorex araneus) // Evolution. Advance online publication 11.10.2011.
См. также:
Ранние этапы видообразования могут сопровождаться множественными геномными перестройками, «Элементы», 02.06.2011.
Варвара Веденина
-
Если бы разное число хромосом принципиально уменьшало бы шансы на скрещивание, любое изменение их числа мгновенно отметалось бы в эволюционном процессе.
-
Не обязательно. Если популяция проходит через "бутылочное горлышко", например, статистическое давление эволюционного отбора не так велико, как вы трактуете.
-
Это, кстати, хорошо обьясняет, почему у бурозубок хромосомные перестройки приводят к образованию хромосомных популяций, а не к мешанине по числу хромосом в пределах одной популяции. Очевидно, хромосомные перестройки получали шанс на закрепление за счёт эффекта основателя.
Правда, обсуждаемая статья путает все карты...
-
-


Ведь трудно представить, чтоб в предковой 24х хромосомной популяции вдруг разом возникли десяток самцов и самок с 23я хромосомами. Скорее, сначала появилась одна "23-самка", которая успешно оплодотворялась "24-самцами" и рожавшая "23-детей". Потом механизм (через много поколений) выключился и изолировал людей от обезьян.
Наверное, можно выделить такие периоды половой изоляции: плодовитые гибриды (собако-волки), стерильные гибриды (мулы и лигры, причем самки гибридов еще могут изредка оплодотворятся самцами исходных видов), утрата механизма развития (оплодотворение поисходит, но зародыш рассасывается), и невозможность оплодотворения.
Последние новости







