Генетическая программа развития пальцев крыла соответствует «динозавровой» теории происхождение птиц

С точки зрения эмбриологии три пальца крыла у птиц соответствуют пальцам II, III и IV исходной пятипалой конечности. Это противоречит палеонтологическим данным, согласно которым у динозавров-теропод и их потомков — ящерохвостых птиц редуцируются пальцы IV и V, а три оставшихся — это пальцы I, II и III. Биологи из Йельского университета измерили активность 14 692 генов в зачатках пальцев у куриного эмбриона и пришли к выводу, что генетическая программа, управляющая развитием первого пальца крыла, соответствует программе развития пальца I, а не II, хотя его зачаток и занимает вторую эмбриональную позицию. Тем самым снимается противоречие, считавшееся важным аргументом против «динозавровой» теории происхождения птиц. Полученный результат подтверждает высказанную в 1999 году гипотезу, согласно которой у динозавров — предков птиц область работы программы формирования пальца I сместилась из первой во вторую эмбриональную позицию.
Так называемая «проблема I-II-III или II-III-IV» уже много десятилетий смущает умы палеонтологов, изучающих происхождение птиц (см. внизу ссылки на статьи ведущего отечественного палеоорнитолога Е. Н. Курочкина).
Палеонтологические находки демонстрируют постепенную утрату пальцев IV и V в эволюционном ряду, ведущем от первых триасовых динозавров к археоптериксу, энанциорнису и другим вымершим ящерохвостым птицам (рис. 2). Значит, если современные (веерохвостые) птицы являются потомками ящерохвостых, то пальцы их крыльев должны быть пальцами I-II-III.
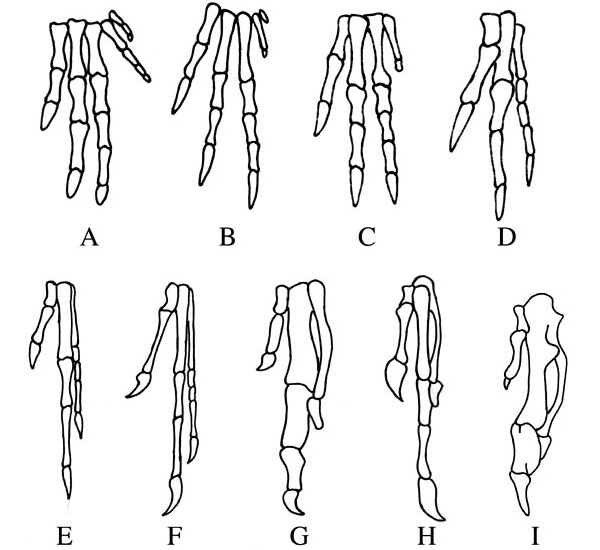
Однако данные эмбриологии этого не подтверждают. Напротив, они указывают, что три пальца птичьего крыла гомологичны пальцам II-III-IV исходной пятипалой конечности наземных позвоночных (рис. 1). В норме у зародышей наземных позвоночных (тетрапод) первым закладывается палец, являющийся продолжением главной оси конечности, и этот палец становится пальцем IV (безымянным). В зачатке птичьего крыла в этой позиции закладывается самый внешний из трех пальцев, то есть третий по счету, если считать от туловища. С точки зрения классической сравнительной эмбриологии, этого вполне достаточно, чтобы считать третий палец крыла гомологичным пальцу IV других тетрапод (и в том числе пальцу IV задних конечностей той же самой птицы). Соответственно, первый и второй пальцы крыла, закладывающиеся с внутренней стороны от третьего, должны быть пальцами II и III.
Но если три пальца крыла у веерохвостых птиц — это действительно пальцы II-III-IV, то тем самым ставится под вопрос их происхождение от ящерохвостых птиц, у которых в крыле сохранились пальцы I-II-III. Это обстоятельство, наряду с другими фактами, заставляет некоторых экспертов предполагать независимое происхождение ящерохвостых и веерохвостых птиц. Если первые уверенно выводятся из динозавров-теропод (в этом сейчас уже никто не сомневается), то предков вторых предлагается искать среди более примитивных триасовых архозавров (см.: Protoavis). Большинство исследователей, однако, убеждены, что веерохвостые птицы произошли от ящерохвостых. С этой точки зрения «проблема I-II-III или II-III-IV» представляется парадоксом, который необходимо разрешить.
Открытие гомеобоксных генов и гомеозисных мутаций пошатнуло классические представления о гомологии, разработанные анатомами и эмбриологами. Действительно, если в результате гомеозисной мутации у дрозофилы вместо антенн вырастают ноги, то чему они гомологичны? Если судить по положению относительно других частей организма (классический сравнительно-анатомический подход), то они, конечно, гомологичны антеннам. Но если судить по генам — регуляторам развития, обеспечивающим формирование этих придатков (то есть по генетической программе их развития), то столь же уверенно можно утверждать, что они гомологичны ногам. С точки зрения сравнительной анатомии это антенны, которые в результате мутации приобрели сходство с ногами, а с точки зрения молекулярной генетики — это ноги, которые в результате мутации переместились в другую эмбриональную позицию. Возможно, правильнее было бы рассматривать обе интерпретации на равных, как две стороны одной медали, но в настоящее время более «модной» становится вторая интерпретация. Именно ей отдают предпочтение авторы обсуждаемой статьи, биологи из Йельского университета. По их мнению, гомология — это «историческая преемственность программы развития, которая определяет онтогенетическую индивидуальность признака».
Ранее уже была высказана гипотеза, согласно которой в эволюции динозавров-теропод имело место гомеозисное событие: «сдвиг рамки» (frame shift), то есть смещение областей работы программ развития пальцев I-II-III на одну эмбриональную позицию в дистальном направлении (от туловища), так что палец I стал развиваться там, где «положено» развиваться пальцу II (G. P. Wagner, J. A. Gauthier. 1999. 1,2,3 = 2,3,4: A solution to the problem of the homology of the digits in the avian hand).

Если эта гипотеза верна, следует ожидать, что генетическая программа развития пальцев крыла должна быть похожа на программу развития пальцев I-II-III других тетрапод (или задних конечностей той же птицы) и отличаться от программы развития пальцев II-III-IV.
Важные аргументы в пользу этого предположения были получены недавно японскими исследователями, которые провели ряд экспериментов по пересадке фрагментов зачатков конечностей у птичьих эмбрионов (Tamura et al., 2011). На внешней стороне зачатка конечностей у наземных позвоночных (там, где закладываются пальцы IV и V) работает «организатор» — группа клеток с включенным геном sonic hedgehog (shh), важнейшим регулятором развития. Оказалось, что если пересадить этот организатор с зачатка крыла (на ранней стадии развития) на внутреннюю сторону зачатка ноги, то там формируются дополнительные пальцы ноги, похожие на палец IV (пальца V у птиц нет ни на крыльях, ни на ногах), но не пальцы крыльев. Если же пересадить организатор с зачатка ноги на внутреннюю сторону зачатка крыла, то там формируются не дубликаты третьего пальца крыла (как можно было бы ожидать, если бы этот палец был гомологичен пальцу IV), а вполне узнаваемые пальцы ноги (они отличаются от пальцев крыла, в частности, по числу фаланг). Кроме того, выяснилось, что третий палец крыла формируется за пределами области экспрессии shh, а не внутри нее, как положено пальцу IV. Эти результаты противоречат предположению о том, что третий палец крыла является пальцем IV.
Биологи из Йельского университета опубликовали в журнале Nature еще более весомые аргументы против теории о соответствии пальцев крыла пальцам II-III-IV. Авторы выделили матричные РНК из зачатков пальцев передних и задних конечностей куриных эмбрионов и определили, к какому гену относится каждая мРНК. Затем по количеству мРНК, считанных с каждого гена, оценили уровень экспрессии этих генов. Всего в зачатках пальцев удалось зарегистрировать активность 14 692 генов. Анализ проводился по отдельности для каждого пальца на двух стадиях развития зачатков («ранней» и «поздней»). Такой глобальный подход — оценка активности всех генов, работу которых удается зарегистрировать в развивающейся конечности, — позволяет ближе всего подойти к той пока еще не поддающейся окончательной расшифровке загадочной сущности, которую обозначают словами «генетическая программа развития».
Выяснилось, что по характеру экспрессии генов первые (самые внутренние) пальцы крыльев и ног четко отличаются от остальных (внешних) пальцев, но при этом они похожи друг на друга. Авторы обнаружили 556 генов, по уровню экспрессии которых можно отличить первый палец (не важно, крыла или ноги) от внешних пальцев. Это весомый аргумент в пользу того, что первый палец крыла — это на самом деле палец I (а не II, как следует из данных эмбриологии).
Очевидно, именно работой этих 556 генов определяется «идентичность» пальцев I с молекулярно-генетической точки зрения. Среди белков, кодируемых этими генами, резко повышена доля регуляторов транскрипции и ДНК-связывающих белков. Это как раз те группы белков, которые и должны, исходя из общих соображений, формировать «программу развития» любых органов.
Внешние пальцы (второй и третий пальцы крыла, второй, третий и четвертый пальцы ноги) сильно отличаются по набору работающих генов от первых пальцев, но вот разобраться в их взаимосвязях друг с другом оказалось трудно. Второй палец крыла похож по своей «генетической программе» отчасти на второй, отчасти на третий палец ноги. Третий палец крыла сочетает черты второго и четвертого пальцев задней конечности.
По-видимому, это означает, что программы развития пальцев в ходе эволюции могут меняться, смещаться из одного эмбрионального положения в другое и даже перекомбинироваться. Полученные результаты свидетельствуют в пользу гипотезы о «сдвиге рамки», потому что первый палец крыла по своей генетической программе явно соответствует первому пальцу ноги, хотя и занимает вторую эмбриональную позицию.
Что касается второго и третьего пальцев крыла, то их программы развития, похоже, сформировались заново из фрагментов программ развития пальцев II, III и IV. Поэтому нельзя определенно сказать, каким пальцам ноги они гомологичны с молекулярно-генетической точки зрения. Последовательное применение того понимания гомологии, которого придерживаются авторы, привело бы к признанию этих пальцев вообще какими-то «новыми» пальцами (и авторы прямо пишут об этом в статье). Но если учесть весь комплекс имеющихся данных (молекулярно-генетическую гомологию первых пальцев, взаимное расположение пальцев крыла, палеонтологические данные и результаты японских исследователей, из которых следует, что третий палец крыла не является пальцем IV), то всё-таки получается, что три пальца птичьего крыла происходят от пальцев I-II-III, а не II-III-IV. А это значит, что конфликт между данными эмбриологии и палеонтологии снят и что эмбриологические данные о развитии пальцев больше нельзя рассматривать как аргумент против «динозавровой» теории происхождения веерохвостых птиц.
Источник: Zhe Wang, Rebecca L. Young, Huiling Xue, Günter P. Wagner. Transcriptomic analysis of avian digits reveals conserved and derived digit identities in birds // Nature. 2011. V. 477. P. 583–586.
См. также:
1) Предки птиц, возможно, были четырехкрылыми, «Элементы», 05.10.2009.
2) Малый размер птичьих геномов — наследие эпохи динозавров, «Элементы», 16.03.2007.
3) В Китае нашли кости гигантского птицеподобного динозавра, «Элементы», 15.06.2007.
4) Как велоцираптор разучился летать, «Элементы», 27.09.2007.
5) Перья сначала служили для красоты, а для полета пригодились позже, «Элементы», 01.11.2008.
6) Е. Н. Курочкин. О четырехкрылом динозавре и происхождении птиц.
7) Е. Н. Курочкин. Новые идеи о происхождении и ранней эволюции птиц.
8) Е. Н. Курочкин. Параллельная эволюция тероподных динозавров и птиц.
-
Мир стал чуточку понятнее. Вариант пальцев опорной конечности не годился для "балансировочных" (на начальном этапе) целей, и произошла генетическая интерференция, преведшая не к редукции (как у тиранозавров), а изменению формы и появлению крыльев. Толковая работа и перевод.
-
: Что касается второго и третьего пальцев крыла, то их программы развития, похоже, сформировались заново из фрагментов программ развития пальцев II, III и IV. Поэтому нельзя определенно сказать, каким пальцам ноги они гомологичны с молекулярно-генетической точки зрения. Последовательное применение того понимания гомологии, которого придерживаются авторы, привело бы к признанию этих пальцев вообще какими-то «новыми» пальцами (и авторы прямо пишут об этом в статье). Но если учесть весь комплекс имеющихся данных (молекулярно-генетическую гомологию первых пальцев, взаимное расположение пальцев крыла, палеонтологические данные и результаты японских исследователей, из которых следует, что третий палец крыла не является пальцем IV), то всё-таки получается, что три пальца птичьего крыла происходят от пальцев I-II-III, а не II-III-IV.
Так. Согласно исследования биологов из Йельского университета, опубикованного в Nature, программы развитися второго и третьего пальцев сформировались из программ развития пальцев II, III и IV, то есть, второй палец крыла точно не является ни пальцем II в целом, ни пальцем III в целом, а третий палец точно не является ни пальцем III в целом, ни пальцем IV в целом. Первый палец является молекулярно-генетически гомологичным пальцу I, но это ни к чему не обязывает второй палец, тем более, что про второй палец точно известно, что его программа развития точно не соответсвует полностью программе развития пальца II. По данным японских исследователей третий палец не является пальцем IV, но это вовсе не обязывает его быть пальцем III, достаточно, чтобы его программа развитися сформировалась не только из программы развития пальца IV, чтобы он не был исключительно пальцем IV. И из этого всего автор новости делает вывод, что второй и третий палец всё-таки являются пальцами II и III, что прямо противоречит выводам биологов Йельского университета, о которых он пишет?????
: А это значит, что конфликт между данными эмбриологии и палеонтологии снят и что эмбриологические данные о развитии пальцев больше нельзя рассматривать как аргумент против «динозавровой» теории происхождения веерохвостых птиц.
Для неискушённого читателя это выглядит так, что был некий спор II-III-IV против I-II-III (если повспоминать публикации на Элементах, то, кажется, связанный со спором о том, от кого произошли современные птицы, выраженный в противостоянии "отечественных" учёных "западным"), в результате исследований выяснилось, что в споре ошибались обе стороны (оказалось что-то вроде I-(II,III)-(III-IV) или даже I-(II,III,IV)-(II,III,IV)), но так уж хочется показать себя правыми, а оппонентов неправыми, что даже считается возможным откровенно... скажем мягко, грешить против логики.
Ничего личного, просто делюсь впечатлениями. Если я где-то ошибся, то буду рад пояснениям.-
Блестящий анализ лишь подтверждающий мудрую, но забытую мысль Мейена: говорить о гомологии и аналогии имеет смысл только определив уровень рассмотрения признаков, систематический вес признаков. Авторы не соблюли этого мудрого правила и в итоге еще больше все запутали - "в споре ошибались обе стороны", то есть ни одна гипотеза de facto не подтверждена и не опровергнута (de jure победу приписали себе любимому). Иного исхода и быть не могло. Любой систематик со времен Линнея понимает, что разные признаки обладают разной значимостью, разным систематическим весом. Поэтому процесс классификации цикличен: сперва классификация организмов, потом классификация признаков, потом уточнение классификации организмов на основе классификации признаков и добавление новых признаков, реклассификация по ним, новое уточнение систематических весов и так далее до насыщения классификации: состояния когда классификация организмов не меняется ни при уточнении старых признаков, ни при добавлении новых. Процедура долгая и утомительная - иной раз на столетия. А хочется быстренько. Поэтому в 70 гг. XX в. придумали нумерологическую систематику (все признаки с равным весом, вопрос решается голосованием), которая с треском показала свою несостоятельность. 40 лет не прошло и снова, значит, на те же грабли - 556 генов. Нужна, как минимум, классификация генов (хотя бы на ортологи и паралоги, или регуляторы первого порядка, второго порядка и т.д.) и сравнение в пределах каждой группы. Сравнивать скопом бессмысленно. Кстати, классификация генов позволяет разрулить ситуацию с дрозофилой с ногами вместо антенн: по регуляторам первого порядка антеннапедии гомологичны ногам, второго - антеннам, а третьего - ни антеннам, ни ногам, так как не обладает ни иннервацией, ни набором рецепторов первых и вторых. И определить по антеннопедиям что возникло в эволюции раньше - ноги или усы - невозможно. Да никто и не пытался. Тем не менее, именно это нам предлагают сделать с пальцами птиц. Так что верно подмечено в комментарии: пошатнулись не классические понятия гомологии и аналогии, разработанные морфологами и эмбриологами, а классические понятия логики (разработанной еще Аристотелем), на которые опирались эти морфологи и эмбриологи, разрабатывая понятия гомологии и аналогии.
-


Последние новости









