Полиплоидные виды вымирают чаще

Американские и канадские ученые оценили скорость появления и вымирания полиплоидных видов растений по сравнению с диплоидными. Их расчеты показали, что полиплоиды менее устойчивы и быстро вымирают; также они оставляют меньше филогенетических потомков по сравнению с диплоидными родственниками. При этом известно, что все семенные растения в своей истории имели полиплоидного предка; известно также, что предок всех цветковых растений на начальных этапах свой эволюции удвоил свой геном и передал его всем потомкам. Таким образом, в кратковременной истории полиплоидия не способствует процветанию видов, зато в редких случаях может оказаться исключительно перспективным приобретением и сработать на долговременную перспективу.
В конце XIX века И. И. Герасимов, ботаник Московского университета, открыл явление полиплоидии — кратного увеличение числа хромосомных наборов. С тех пор интерес к этому явлению остается исключительно высоким как у академиков, так и у практиков. Селекционеры используют полиплоидию для выведения новых сортов культурных растений (множество культурных растений полиплоидны: пшеница, ячмень, кофе, банан, сахарный тростник, табак, слива, клубника, шпинат, укроп, яблоня, редиска, виноград), дрожжей и т. д. Для фундаментальной науки важнейшим является вопрос, какую роль играла (и играет) полиплоидия в эволюционном процессе. На этот счет приводятся прямо противоположные мнения, например вот такие:
...polyploidy has contributed little to progressive evolution (Stebbins, 1971) («...полиплоидия почти ничего не привнесла в поступательный эволюционный процесс»);
....polyploidy, far from playing a secondary role in evolution, has provided the additional, uncommitted gene loci necessary for major steps in the evolution of animals. (Schultz, 1980) («...полиплоидия оказалась далеко не последним инструментом в эволюции животных, обеспечив геном не связанными ничем локусами, столь необходимыми для важных эволюционных подвижек»).
Сейчас эти два мнения сосуществуют, хотя второму всё чаще отдается предпочтение. Так, современные ботаники признают, что большинство, если не все, сосудистые растения в своей прошлой филогенетической истории имели полиплоидных предков. Это не так-то просто обнаружить, потому что геномы полиплоидов эволюционируют, изменяя до неузнаваемости полученные копии.
Однако современные методы геномной статистики позволяют выявить такие древние скачки в плоидности предковых организмов. Для цветковых растений, например, это с успехом проделала группа ученых под руководством Клода де Памфилиса (Claude dePamphilis) из Университет штата Пенсильвания.
Чтобы подсчитать число вероятных событий полиплоидии, ученые использовали данные по последовательностям у видов цветковых растений с полностью прочтенными геномами, расположив генные гомологи на временной оси согласно меткам молекулярных часов. Выяснилось, что у всех цветковых растений имеются следы двух событий полиплоидизации: первое испытал предок всех семенных растений, второе — предок всех цветковых растений. На верхнем рисунке эти события выделяются двумя пиками частоты полиплоидных узлов относительно временной шкалы.

Существенно проще и надежнее подсчитать число недавних (относительно недавних, естественно) событий полиплоидии — тех, которые произошли в пределах родовых групп. Иными словами, выявить те виды растений, у которых геном удвоился или утроился (учетверился и т. д.) уже после появления предка всего рода. По современным оценкам, около 35% видов ныне живущих сосудистых растений имеют полиплоидный геном, сформировавшийся уже после зарождения рода (в англоязычной литературе они получили название neoploids, или «неоплоиды» в буквальном переводе). Столь высокие оценки числа новых полиплоидов не означают, однако, что полиплоидия влечет быстрое видообразование. Возможно, что полиплоиды просто часто появляются в ходе эволюции.
В этих альтернативных гипотезах попытались разобраться биологи из Университета Британской Колумбии (Ванкувер, Канада) и Университета Дьюка (Дарем, США), Аризонского (Тусон, США) и Индианского (Блумингтон, США) университетов. Они сравнили скорости диверсификации, видообразования и вымирания среди диплоидных и полиплоидных представителей родов. Все расчеты основывались на филогенетических деревьях тех родов, для которых, во-первых, имеются достаточно представительные и выверенные данные по видовому составу и видов в которых не менее десятка; во-вторых, имеются данные по геномике, а не только по морфологической эволюции (иначе статистические деревья нельзя было бы составить); в-третьих, найдены данные по числу хромосом у не менее чем 65% видов рода; в-четвертых, в составе рода должен быть хотя бы один полиплоид.
В доступных базах данных набралось 63 рода, удовлетворяющих всем условиям. Из них 49 родов цветковых растений и 14 родов папоротников. В каждом роде в среднем насчитывалось по 39 видов с известным числом хромосом. Это число покрывает 60–70% видового разнообразия данных родов. С ними-то ученые и работали. Для каждого выбранного рода они составили деревья, на которых отметили полиплоидов — так, как это показано на нижнем рисунке.
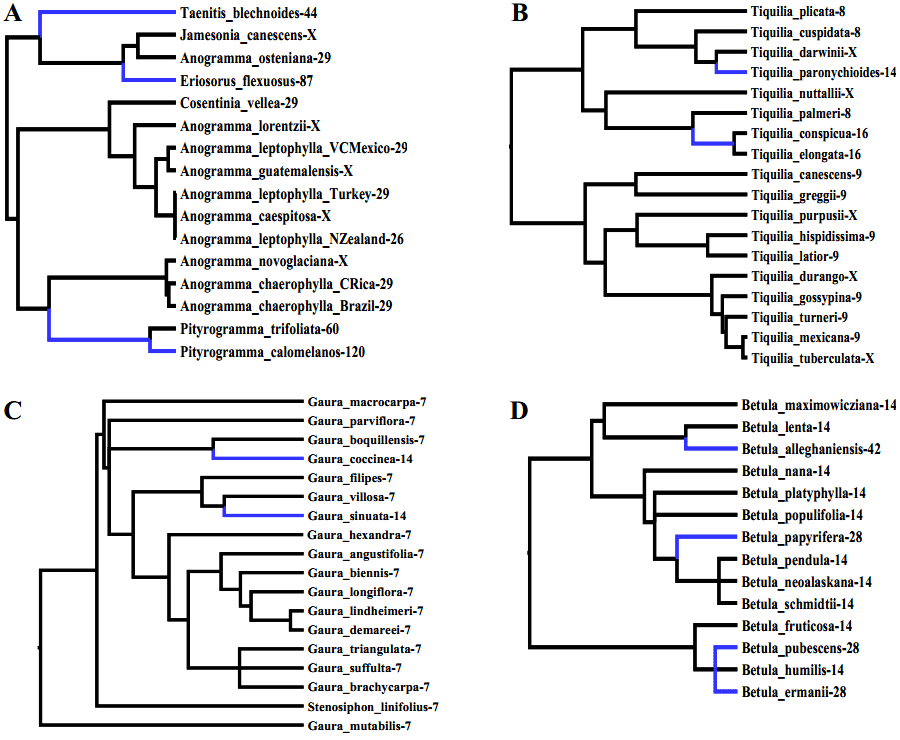
Анализируя все 63 схемы, ученые выделили важную особенность: подавляющее большинство полиплоидных веточек располагаются на концах филогенетических деревьев, и очень редко синие (полиплоидные) ветви имеют сколько-нибудь значительную длину. Из этого неизбежно следует, что полиплоиды существуют недолго, дают мало эволюционных потомков и быстро вымирают.
Проверить данное следствие, хотя оно вполне закономерно увязывается с приведенными схемами, ученые смогли с помощью сложной статистики. Они рассчитывали скорости видообразования, вымирания и диверсификации. Последняя величина связана с вымиранием и видообразованием и является в некоторой степени производной от них. Темпы видообразования у диплоидных видов были явно выше, чем у полиплоидов, о чём свидетельствуют высокие вероятности более активного видообразования у диплоидов у большинства родов (графики А и В); при этом вероятности вымирания у диплоидных видов ниже, чем у полиплоидов. Таким образом, полиплоидные виды появляются относительно часто, но зато оказываются недолговечными и быстро вымирают.
Отмечу, что в палеонтологической летописи морских беспозвоночных подобных эфемерных видов (появляющихся и, не оставив потомков, быстро вымирающих) регистрируется сравнительно много — около 45% от текущего разнообразия. Это количество оставалось более или менее постоянным на протяжении фанерозойской истории. Какую роль играла в формировании эволюционных эфемеров полиплоидия, еще предстоит выяснить.
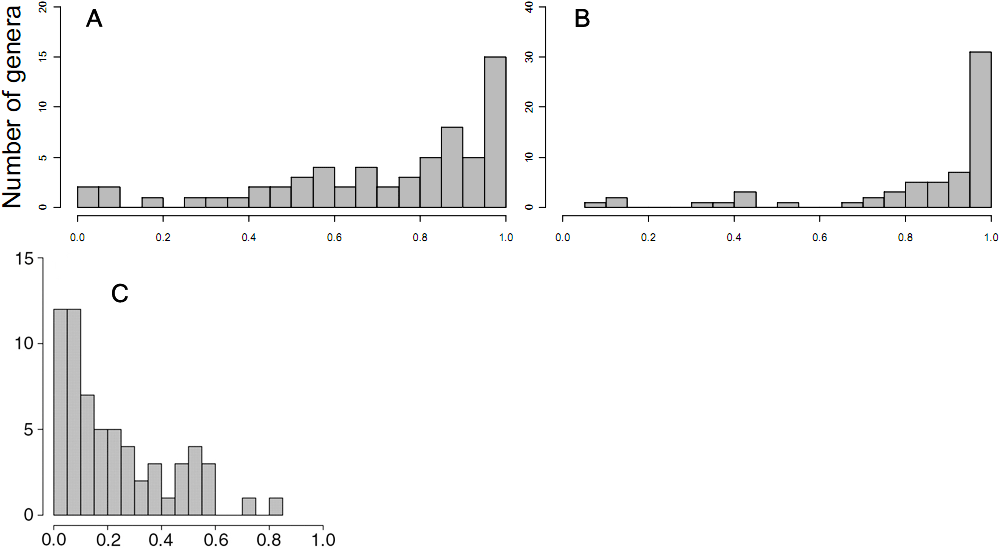
Из этих расчетов складывается представление о долговременных и кратковременных эффектах полиплоидии в эволюции растений. В кратковременном (опять же — в геологическом, эволюционном) масштабе полиплоидия никак не способствует устойчивости видов и не вызывает всплесков видообразования. Однако некоторые, весьма редкие, полиплоидные попытки оказываются необычайно ценными и своевременными, и тогда на базе обновленного удвоенного генома формируются прогрессивные зачинатели целых семейств, классов или даже типов.
Источники:
1) Yuannian Jiao, Norman J. Wickett, Saravanaraj Ayyampalayam, André S. Chanderbali, Lena Landherr, Paula E. Ralph, Lynn P. Tomsho,Yi Hu, Haiying Liang, Pamela S. Soltis, Douglas E. Soltis, Sandra W. Clifton, Scott E. Schlarbaum, Stephan C. Schuster, Hong Ma, Jim Leebens-Mack, Claude W. dePamphilis. Ancestral polyploidy in seed plants and angiosperms // Nature. 2011. V. 473. P. 97–100.
2) Itay Mayrose, Shing H. Zhan, Carl J. Rothfels, Karen Magnuson-Ford, Michael S. Barker, Loren H. Rieseberg, Sarah P. Otto. Recently Formed Polyploid Plants Diversify at Lower Rates // Science. 2011. V. 333. P. 1257.
Елена Наймарк
-
Логично получается. Полиплойдный геном тормозить изменчивость. Мутации, возникшие в одной из пар хромосом блокируются или нивелируются аналогичными немутантными генами в других хромосомах. В результате растение медленнее приспосабливается к новым условиям обитания и проигрывают в конкурентной борьбе.
-
Нет, не тормозит. Полиплоидность не дает проявиться отрицательным мутация, поскольку нужный белок будет производиться немутированным геном-дупликатом. Но зато, во второй копии генома могут происходить одна за другой мутации, которые могут привести к образованию белков с полезными новыми свойствами. Трудно ожидать, что новое полезное свойство разовьется в результате одной мутации. Например, для появления у приматов гена белка, чувствительного к красному цвету (из белка, чуствительного к зеленому свету) понадоболось 5 мутаций. Зато приматы в результате обрели богатое цветовое зрение. Многие белки, являющиеся гомологами легкой цепи иммуноглобулинов, скорее всего появились по такому же механизму. Hе мутированный ген работал себе как надо, а в это время в геноме сидел параллельный ген, постепенно мутирующий к новому качеству.
Так что, удвоение генов - основа эволюции. Позволяет накапливаться положительным мутациям, защищая организм от гибели в результате появления "испорченного" белка.-
Вероятно, я неверно сформулировал. На скорость мутаций полиплодия скорее всего не влияет, а мутация в одном из экземпляров хромосомы с некоторой долей вероятности (как Вы справедливо заметили) окажется рецессивной и будет экспрессироваться правильный белок из немутантной хромосомы. Таким образом, не смотря на некоторые генетические изменения, фактически фенотипических изменений не будет. И так будет происходить как с вредными, так и с полезными мутациями, поэтому в целом полиплойдный вид будет медленней приспосабливаться к изменениям в окружающей среде.
-
-


-
Рассуждение вполне логично. Однако нужно иметь в виду, что, во-первых, северная (бореальная, арктическая?) среда не обязательно менее стабильна. Она скорее дальше от абиотического оптимума для большей части растений. Во-вторых, есть ли у Вас данные о доле полиплоидов в северных и тропических регионах - просто для сравнения, чтобы понимать, обсуждаем ли мы фантазии или факты. В-третьих, процент аллополиплоидов совсем невысок, авторы статьи приводят эту цифру (я забыла, какая точно). В четвертых, распространение новых полиплоидов по климатическим зонам и ландшафтам наверняка исследовано, это очень "горячая" тема в ботанике. Если найдете что-то интересное на эту тему, дайте знать.
-
Бореальная и арктическая среда менее стабильна в том отношении, что она недавно была полностью (или почти полностью) очищена от растений; в прошлом это случалось неоднократно на довольно коротком временном интервале. В тропиках такого все-таки не было.
Доля полиплоидов действительно увеличивается в высоких широтах - это указано во многих статьях и книгах, хотя выполняется,видимо, не для всех групп, а только для флоры в целом (и не для всех регионов, а там, где с широтой действительно четко меняется климат).
Может быть, процент аллополиплоидов невысок во всей флоре; но каков этот процент от общего числа полиплоидов на севере? По крайней мере для некоторых групп он довольно высок(http://www.amjbot.org/content/97/2/324.full; http://www.ncbi.nlm.nih.gov/pubmed/19120990). Первая ссылка - на статью 2010 г про однолетние злаки рода пукцинелла (бескильницы). Вторая ссылка - на тоже довольно свежую (2008) статью про пальцекорники. В обоих случаях по крайней мере часть аллополиплоидов возникла "на месте", а не мигрировала с юга.
Насколько я знаю, проблемой гибридизации растений и в частности образованием полиплоидов на севере Европы интересуется А.Б. Шипунов (думаю, он знает много интересного на эту тему). Судя по его рассказам, гибридизация на севере Европы действительно происходит чаще, чем на юге (чаще ли при этом образуются полиплоиды - не знаю).
-
Последние новости








