Арнеболагус — звезда первой величины в эволюции зайцеобразных
Алексей Лопатин, академик, Палеонтологический институт имени А. А. Борисяка РАН (Москва),
Александр Аверьянов, доктор биологических наук, Зоологический институт РАН (Санкт-Петербург)
«Природа» №2, 2021

Обычно мы не осознаем, насколько удивительны некоторые вроде бы хорошо знакомые нам животные. Вот и зайцы с кроликами, а также менее известные пищухи оказываются довольно странными созданиями, если смотреть на них глазами зоолога. Во-первых, сильно удлиненные задние конечности они используют не для двуногого передвижения прыжками, как кенгуру или тушканчики, а для быстрого четвероногого галопа. Во-вторых, только у зайцеобразных подошвы лап полностью покрыты густой шерстью. Детали строения их зубов очень сложно сопоставить с бугорками и складками на зубах других плацентарных млекопитающих. Боковые стороны костей передней части черепа зайцеобразных перфорированы множеством отверстий разной формы и величины, в результате чего они имеют вид ажурной решетки, не имеющей аналогов у других млекопитающих. Костное нёбо очень сильно сокращено, а пяточная кость пронизана диагональным каналом, которого нет у других зверей [1]. Желудок состоит из двух функциональных отделов: в переднем происходит бактериальное брожение, а в заднем — переваривание пищи. Есть у зайцеобразных и другие особенности, ставшие причиной для разногласий при оценке их родственных связей с другими млекопитающими.
Общим обликом, отсутствием клыков и наличием постоянно растущих крупных резцов с большим промежутком за ними (диастемой) зайцеобразные напоминают грызунов, что долгое время позволяло систематикам рассматривать две эти группы в составе единого отряда Rodentia. В 1912 г. американский палеонтолог и зоолог Дж. Гидли [2] обосновал принадлежность зайцеобразных к особому отряду Lagomorpha и выдвинул гипотезу об их близком родстве с парнокопытными, основанную на поверхностном сходстве с некоторыми древними формами Artiodactyla и общем поперечном способе жевания.
Увлекшись идеей об отсутствии ныне живущих родичей зайцеобразных, некоторые систематики в начале XX в. полагали, что их можно напрямую связывать с мезозойскими группами, вплоть до триконодонтов [3] (правда, о млекопитающих мезозоя тогда было известно очень мало). Несколько позже особенности репродуктивной системы зайцеобразных убедили некоторых зоологов в близком родстве Lagomorpha с сумчатыми [4]. Очевидно, эти гипотезы теперь имеют значение лишь для историков науки. Но даже в конце XX в. известный американский специалист по млекопитающим М. Маккенна [5] развивал поддержанную многими исследователями идею о том, что зайцеобразные — это остаток некогда процветавшей особой древней ветви плацентарных, названной им анагалидами [6]. Самыми ранними представителями анагалид он признавал позднемеловых заламбдалестид — вымерших млекопитающих, предположительно способных к четвероногому прыжковому передвижению. Общей чертой анагалид считались высококоронковые зубы.
Была также предпринята попытка (основанная на изучении белковых последовательностей) связать зайцеобразных с приматами и тупайями [7]. Но современные молекулярно-генетические данные вновь заставляют объединять зайцеобразных с грызунами — на этот раз в надотрядной группе грызунообразных (Glires, или Gliriformes), которая вместе с приматами, шерстокрылами и тупайями включается в состав большей группировки Euarchontoglires [8].
В палеоцене и эоцене Азии найдены многочисленные остатки миксодонтов (Mixodontia) — группы, предковой для зайцеобразных (теперь они вместе объединяются в ветвь двупарнорезцовых — Duplicidentata). По строению зубной системы миксодонты напоминают одновременно и зайцеобразных, и грызунов (поэтому они и получили свое латинское название, означающее смешаннозубые), но не имеют отношения к происхождению последних. Среди миксодонтов по строению черепа и зубов наибольшую близость к зайцеобразным обнаруживает группа мимотонид (Mimotonida). Для их общей ветви устоялось странное по звучанию, но логичное по содержанию название зайцеобразнообразные (Lagomorphamorpha).
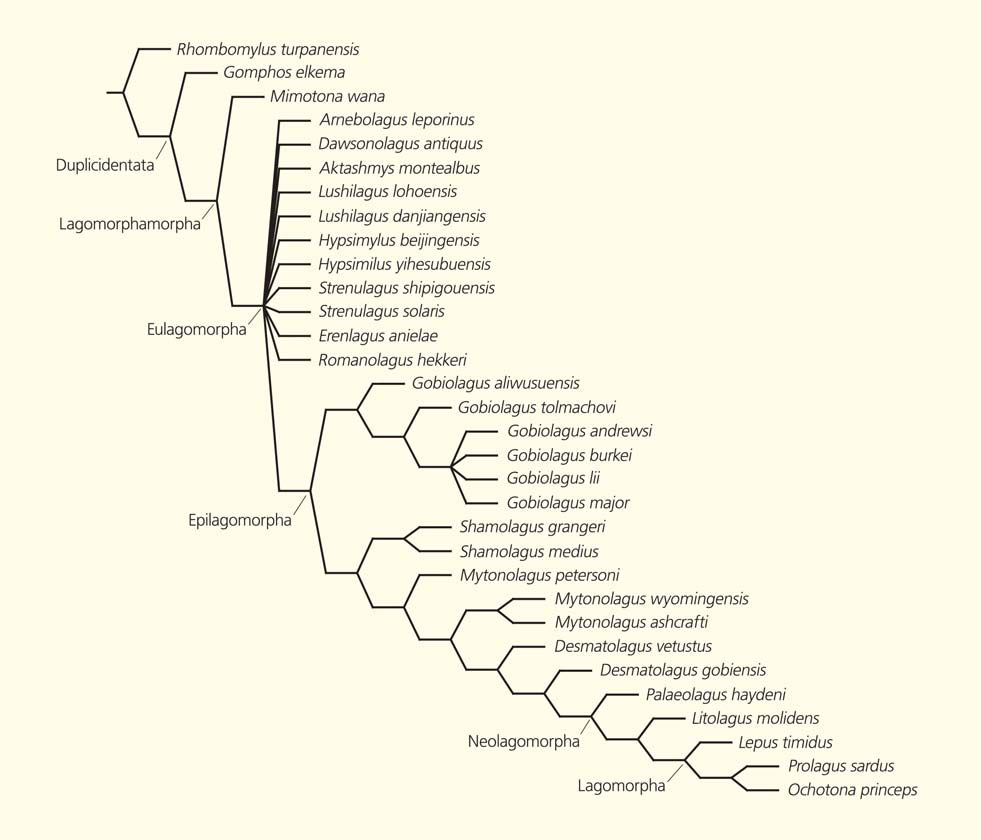
Филогенетическое древо двупарнорезцовых [10, fig. 4]

Реконструкция представителя миксодонтов. Фигурка из мамонтового бивня, вырезанная М. Дашцэрэном по рисунку А. А. Гуреева. Фото П. Ю. Пархаева
В конце палеоцена (56–55 млн лет назад) разнообразные миксодонты (включая мимотонид и больше похожих на грызунов эвримилид) доминировали в азиатских сообществах среди мелких растительноядных млекопитающих, а со второй половины раннего эоцена (около 50 млн лет назад) с ними сосуществовали довольно многочисленные архаичные зайцеобразные. По-видимому, ответвление зайцеобразных от мимотонид произошло на рубеже этих двух геологических эпох, когда в Азии впервые появились некоторые другие группы растительноядных млекопитающих, в том числе парнокопытные и непарнокопытные.
Древнейший представитель зайцеобразных — Arnebolagus leporinus, описанный в 2008 г. из базального эоцена Монголии [9], имеет возраст около 55 млн лет. Родовое название ему было дано в честь звезды Арнеб — α созвездия Зайца. Оно подчеркивает древность и первостепенную важность этой формы для изучения эволюции группы. Арнеболагус был найден в местонахождении Цаган-Хушу, расположенном во впадине Нэмэгт в Южной Гоби, в пачке бумбан свиты наран-булак. Первоначально арнеболагус был описан по единственному верхнему предкоренному зубу, что, конечно, затрудняло оценку его положения на эволюционном древе зайцеобразных и мимотонид. Изучение дополнительных материалов (верхние предкоренные и коренные зубы и один нижний коренной) позволило нам недавно более детально охарактеризовать эту древнюю форму и с помощью анализа 54 морфологических признаков 32 видов реконструировать родственные связи ранних зайцеобразных [10].
В результате мы предложили считать арнеболагуса основателем новой ветви зайцеобразных современного типа, получившей название эулагоморф (Eulagomorpha, т.е. настоящие зайцеобразные). К ней относятся все зайцеобразнообразные, кроме мимотонид. Важнейшая характеристика эулагоморф — верхние щечные зубы односторонне-высококоронкового строения (внутренняя сторона коронки по высоте примерно вдвое превышает наружную), как у арнеболагуса. Для конца раннего эоцена (около 50–48 млн лет назад) известно не менее семи родов эулагоморф из разных частей Азии, включая современные территории Индии, Киргизии, Китая и Монголии.

Верхние щечные зубы и реконструкция правого зубного ряда Arnebolagus leporinus. P3 — предпоследний верхний премоляр, P4 — последний верхний премоляр (на реконструкции отображен зеркально), M1 — первый верхний моляр, M2 — второй верхний моляр [10, fig. 1, 2 с изменениями]
Внутри эулагоморф около 45 млн лет назад обособилась ветвь, названная эпилагоморфами (Epilagomorpha, т.е. высшие зайцеобразные), которая стала предковой для всех более поздних зайцеобразных, включая несколько вымерших семейств и современных зайцев, кроликов и пищух. В начале среднего эоцена (около 40 млн лет назад) эпилагоморфы распространились в Северную Америку, где дали начало ряду местных форм.
Собственно зайцеобразные (Lagomorpha) в объеме современных семейств заячьих и пищуховых и их ближайших общих предков возникли приблизительно на рубеже эоцена и олигоцена (около 35 млн лет назад), причем заячьи (Leporidae) сформировались в Северной Америке, а пищухи (Ochotonidae) — в Азии. Позже зайцы и кролики проникли в Европу, Африку и Южную Америку. В историческое время человек помог им расселиться в Австралии и на многих островах. Среди зайцеобразных всегда преобладали сравнительно мелкие и легко сложенные формы. Лишь островной гигантский заяц Nuralagus rex (он жил 5–3 миллиона лет назад на объединенных в те времена Балеарских о-вах), не имевший хищных врагов, достигал веса более 20 кг и отличался массивным обликом [11]. Необходимо отметить, что известное уже сегодня разнообразие родов и видов вымерших зайцеобразных в несколько раз превосходит их современное разнообразие.
Помимо арнеболагуса, комплекс мелких млекопитающих из пачки бумбан местонахождения Цаган-Хушу включал многочисленных миксодонтов, грызунов и насекомоядных [12], а также редких приматов. Сравнение с примерно одновозрастными фаунистическими комплексами Китая, в которых весьма обильно представлены лесные формы, позволяет предположить, что в начале эоцена в районе впадины Нэмэгт Южной Гоби преобладали открытые ландшафты. Именно такие местообитания благоприятствовали распространению древнейших эулагоморф — арнеболагусов.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (проект 19-04-00057).
Литература
1. Bleefeld A. R., Bock W. J. Unique morphology of lagomorph calcaneus // Acta Palaeontologica Polonica. 2002; 47(1): 181–183.
2. Gidley J. W. The lagomorphs an independent order // Science. 1912; 36(922): 285–286.
3. Gidley J. W. Evidence bearing on tooth-cusp development // Proceedings of the Washington Academy of Science. 1906; 8: 91–110.
4. Petrides G. A. A fundamental sex difference between lagomorphs and other placental mammals // Evolution. 1950; 4(1): 99.
5. McKenna M. C. Early relatives of Flopsy, Mopsy, and Cottontail // Natural History. 1994; 103(4): 56–58.
6. McKenna M. C., Bell S. K. Classification of mammals: above the species level. N.Y., 1997.
7. Graur D., Duret L., Guoy M. Phylogenetic position of the order Lagomorpha (rabbits, hares and allies) // Nature. 1996; 379(6563): 333–335.
8. Murphy W. J., Eizirik E., O’Brien S. et al. Resolution of the early placental mammal radiation using Bayesian phylogenetics // Science. 2001; 294(5550): 2348–2351.
9. Лопатин А. В., Аверьянов А. О. Древнейшее зайцеобразное (Lagomorpha, Mammalia) из базального эоцена Монголии // Доклады Академии наук. 2008; 419(5): 709–711.
10. Lopatin A. V., Averianov A. O. Arnebolagus, the oldest eulagomorph, and phylogenetic relationships within the Eocene Eulagomorpha, new clade (Mammalia, Duplicidentata) // Journal of Paleontology. 2020: 1–12. DOI: 10.1017/jpa.2020.94.
11. Quintana J., Kohler M., Moya-Sola S. Nuralagus rex, gen. et sp. nov., an endemic insular giant rabbit from the Neogene of Minorca // Journal of Vertebrate Paleontology. 2011; 31(2): 231–240. DOI: 10.1080/02724634.2011.550367.
12. Lopatin A. V. Early Paleogene insectivore mammals of Asia and establishment of the major groups of Insectivora // Paleontological Journal. 2006; 40 (Suppl. 3): S205–S405. DOI: 10.1134/S0031030106090012.
-
Здравствуйте, хочу поделиться с вами таким интернет-магазином как автоматика для дома, тут вы сможете найти не только всё для ворот и автоматики , а еще и все для дома/дачи, перил, каркасов и многое другое, в общем кому интересно, всем туда - https://www.1020.by/katalog/otkatnye-vorota-pod-klyuch.html


Избранное


См. также









Художественная реконструкция арнеболагусов в среде их обитания. Рисунок А. А. Атучина