Расцвет и закат гетероспоровых плауновидных
Сергей Владимирович Наугольных,
доктор геолого-минералогических наук, Геологический институт РАН (Москва, Россия)
«Природа» №7, 2019

Плауновидные — очень интересная группа растений, стоящая несколько особняком в современном растительном мире. Их рассматривают как один из типичных примеров «живых ископаемых», т.е. организмов сохранившихся с далекого прошлого до современности.
В большинстве наиболее авторитетных ботанических руководств плауновидные рассматриваются в качестве самостоятельного отдела Lycopodiophyta. К этой группе растений в современном мире относятся вечнозеленые травянистые или полукустарниковые формы. В минувшие геологические эпохи, в особенности в позднем палеозое, среди плауновидных было много крупных древовидных растений, но об этом чуть позже. Листья плауновидных (филлоиды1) простые, с единственной средней жилкой, а спорофиллы (фертильные «листья» со спорангиями) собраны в стробилы, или фертильные зоны. Для плауновидных, как и для многих других растений, характерно чередование поколений: с гаплоидным (гаметофиты) и с диплоидным набором хромосом (спорофиты). Гаметофиты плауновидных довольно разнообразны. Они могут быть однополыми или обоеполыми, способны произрастать в почве или прямо на поверхности субстрата. Мужские гаметофиты продуцируют спермии, которые после оплодотворения женского гаметофита дают начало спорофиту — крупному растению, продуцирующему спорангии и, соответственно, споры.
У гомоспоровых плауновидных споры примерно одинакового размера и формы. У гетероспоровых плауновидных споры двух разных типов — мелкие (микроспоры; со средним диаметром около 30–50 мкм) и крупные (мегаспоры, со средним диаметром около 400–600 мкм). Из микроспор после их попадания в субстрат образуются мужские гаметофиты, а из мегаспор — женские. Таким образом, споры, прорастая, в свою очередь дают начало следующему поколению гаметофитов. Циклы повторяются, сменяя друг друга. Свою геологическую историю плауновидные ведут с позднего силура, т.е. это одна из древнейших групп высших растений2.

Современные гомоспоровые плауновидные Lycopodium annotinum: заросли и отдельное растение. Здесь и далее длина масштабной линейки 1 см
Рассказ о необычной и в чем-то даже поучительной истории расцвета и заката гетероспоровых плауновидных хочется начать со следующего. Эти незаметные растения сопровождают нас практически постоянно — и в дни радостей, и в дни лишений. Так, на полях Великой Отечественной в качестве подсушивающей присыпки на раны использовали споры плауна ликоподиума.

Современное гетероспоровое плауновидное Isoetes lacustris
Среди современной растительности плауновидные занимают весьма скромное положение. Они представлены несколькими родами, к которым относятся исключительно небольшие травянистые или полукустарничковые формы. Роды распределены по двум большим группам, таксономический статус которых разными ботаниками понимается несколько по-разному. По моему твердому убеждению, эти группы должны иметь ранг никак не ниже класса, хотя обе они (и, соответственно, оба этих класса) принадлежат к одному отделу плауновидных.
К первому классу (Lycopodiopsida) относятся гомоспоровые (иначе говоря, равноспоровые, или изоспоровые) плауновидные, со спорами примерно одинаковой формы и размера. Ко второму классу (Isoetopsida) относятся гетероспоровые (или разноспоровые) плауновидные, у которых есть как маленькие споры (они созревают в микроспорангиях), так и крупные (они созревают в мегаспорангиях). При этом форма и размер самих микро- и мегаспорангиев могут практически не различаться.
Из гомоспоровых плауновидных в первую очередь надо назвать известный всем любителям ботаники род ликоподиум (Lycopodium) и близкие ему роды Huperzia и Diphasiastrum, самостоятельность которых признается не всеми ботаниками. Из гетероспоровых плауновидных до современности дожили настоящие «живые ископаемые» — шильник, или полушник-изоэтес (Isoetes), который встречается и в России, а также стилитес (Stylites), обнаруженный сравнительно недавно в Южной Америке, в Андах, где он произрастает на хорошо увлажняемых высокогорных альпийских лугах. Кроме этого, к гетероспоровым относится селагинелла (Selaginella), внешне напоминающая ликоподиум. Но спорангии у селагинеллы, как правило, собраны не в терминальный стробил3, а в фертильную зону с активной апикальной меристемой4, т.е. со способностью побега пролиферировать, расти дальше над фертильной зоной после созревания спор. Для всех гетероспоровых плауновидных характерна одна важная особенность. У них есть язычок, или лигула — специальный орган, расположенный либо в пазухе филлоида, либо дистальнее5 спорангия на адаксиальной стороне (т.е. обращенной к оси побега) спорофилла.
Перечисленными выше растениями практически исчерпывается разнообразие современных плауновидных. При этом подавляющее большинство ныне живущих представителей этой некогда процветавшей группы относится к гомоспоровым плауновидным семейства Lycopodiaceae. Гетероспоровые плауновидные гораздо менее заметны и не столь многочисленны.
Заглянем в прошлое
Однако в минувшие геологические эпохи ситуация была обратной. В растительности позднего палеозоя и раннего мезозоя доминировали гетероспоровые плауновидные, и представлены они были преимущественно не травянистыми, а древовидными формами. Именно к этой группе принадлежали гигантские лепидодендроны, сигиллярии и ботродендроны, произраставшие на Земле в каменноугольном периоде и образовывавшие знаменитые леса карбона, остатки которых в виде каменного угля так помогли человечеству на пути технологического прогресса. Лепидодендроны и их родственники представляли собой огромные деревья высотой до 30 м, а иногда и более, с широкой раскидистой кроной. Мне довелось видеть гигантский окаменелый пень лепидодендрона, хранящийся в музее г. Оснабрюке (Северный Рейн — Вестфалия, Германия). Диаметр пня почти 2 м! От пня в стороны расходились ризофоры — корневые поддержки (не корни!), к которым прикреплялись длинные и тонкие цилиндрические аппендиксы, которые и были настоящими корнями лепидодендронов и их родственников. Ризофоры лепидодендронов и сигиллярий относятся к особому формальному роду стигмария (Stigmaria). Здесь уместно добавить, что в силу специфики палеоботаники различные органы одного и того же растения нередко относят к разным формальным родам6.
В России также известны весьма представительные местонахождения ископаемых остатков гигантских древовидных каменноугольных плауновидных. Из наиболее знаменитых можно назвать обнажения песчаников и алевролитов с углистыми прослоями, расположенные по берегам р. Северский Донец в Ростовской обл. Здесь встречаются стволы лепидодендронов (а также их листья и стробилы) вместе с многочисленными ископаемыми остатками других растений, свидетельствующими о буйстве жизни в каменноугольных лесах и болотах. Именно подобные растительные остатки и послужили материнским веществом для каменных углей Донбасса.
Во время Великой Отечественной войны, когда Донбасс был оккупирован, уголь в топки наших бронепоездов поступал с Урала, из Кизеловского угольного бассейна. Здесь, так же как и в Донбассе, в мощных слоях песчаников, содержащих угольные пласты, можно найти фрагменты коры и даже целые стволы лепидодендронов. Последние исследования показали, что каменноугольные отложения Кизеловского бассейна могут дать еще много интересных палеоботанических находок [1].
И, что наверняка удивит многих читателей, остатки древовидных плауновидных каменноугольного периода можно найти совсем недалеко от Москвы — в Тверской, Тульской и Калужской областях. Более того, крупный стробил гетероспорового древовидного плауновидного, отнесенный к новому роду и виду Moscvostrobus mirabilis, был найден в пределах Московской обл., в карьере неподалеку от г. Серпухова [2].
Таким образом, тема гетероспоровых плауновидных тесно соприкасается с насущными нуждами человечества, пусть даже и не всегда явным образом.
Если обратиться к учебникам ботаники, причем не только к школьным, но и к вузовским, то в них мы найдем обобщенное описание эволюции плауновидных, согласно которому когда-то эти растения были большими, а потом постепенно, за счет общей соматической редукции, стали уменьшаться в размерах, чуть ли не деградировать. И в итоге эволюции превратились в современные маленькие плауны. По сути своей эта картина очень упрощена, более того, она даже неверна. Эволюционная история представляется иной, особенно если оценивать ее, анализируя конкретные палеоботанические факты.
Что выяснилось
Впервые всерьез проблемой эволюции гетероспоровых плауновидных занялся немецкий палеоботаник К. Мэгдефрау [3, 4]. Именно его концепция широко вошла в палеоботаническую литературу. Исследователь считал, что от каменноугольных гетероспоровых плауновидных (в частности, от рода сигиллярия) вследствие постепенной редукции в ходе эволюции произошли плевромейевые (семейство Pleuromeiaceae с типовым родом Pleuromeia), т.е. гетероспоровые плауновидные, широко распространившиеся практически по всей Земле в начале триасового периода. Плевромейевые, в отличие от своих каменноугольных предков, уже не были такими большими растениями. Их побеги не ветвились. На верхушке побега располагался единственный стробил (у некоторых видов он обладал пролиферацией7 и, таким образом, представлял собой фертильную зону8). Не было у плевромейевых и широко ветвящихся стигмариеподобных ризофоров. Вместо них развивался похожий на якорь клубневидный ризофор, иногда рассеченный на продольные лопасти. По мнению Мэгдефрау, плевромейевые, в свою очередь, тоже испытали общую редукцию, и к началу мелового периода они превратились в формы с укороченными (высотой до 20 см) побегами, как у представителей рода натгорстиана (Nathorstiana), описанного в 1932 г. Мэгдефрау из нижнемеловых отложений Германии, которые обнажаются в местечке Кведлинбург в Гарце. Мэгдефрау предположил, что морфотип натгорстианы в процессе дальнейшей эволюции испытал редукцию и дал план строения современных шильников. У этих современных растений побеги укороченные. По существу, они превращены в запасающий орган с небольшим расширением в основании, напоминающим ризофор плевромейи и натгорстианы. Но иногда побег изоэтеса ветвится, а в исключительных случаях ветвится даже повторно, что может быть истолковано как атавистический признак — повторение строения ветвящихся крон гигантских каменноугольных плауновидных [5].
Такой построенный им эволюционный ряд — от гигантских каменноугольных лепидодендронов до современного шильника, — выразившийся в перманентной соматической редукции спорофита этих растений, выглядел очень логично и был подтвержден хорошо задокументированными палеоботаническими находками.
Однако по мере появления новых данных филогенетическая схема, предложенная Мэгдефрау, начала меняться. Так часто бывает и в палеоботанике, и в других науках, когда новые факты вступают в противоречие с общепринятыми концепциями.
На Урале, а именно, в Суксунском р-не Пермского края, на левом берегу живописной р. Сылвы располагается широко известное среди палеонтологов и стратиграфов местонахождение Чекарда. Оно знаменито тем, что здесь встречаются ископаемые остатки растений и насекомых раннепермского возраста, причем окаменелости отличаются просто изумительной, уникальной сохранностью [6, 7].
Мне тоже выпало счастье принять участие в изучении и описании палеонтологических сокровищ Чекарды. В плитчатых алевролитах, аргиллитах и мергелях сохранились практически все основные представители растительного мира пермского периода, характерные для флор Северного полушария.
Среди остатков растений, собранных в этом местонахождении, обнаружился один очень странный и необычный отпечаток. Как выяснилось в процессе изучения, это спорофилл плауновидного, но его морфология была совершенно нетипичной для палеозойских представителей данной группы. Он не был похож на спорофиллы лепидодендронов и сигиллярий, но в гораздо большей степени напоминал спорофиллы плевромейевых.
Однако в ранней перми (а отложения именно такого возраста, как уже было отмечено, обнажаются в Чекарде) никаких плевромейевых, по существовавшим на тот момент представлениям, быть не должно.
Это новое растение было описано в качестве нового рода и вида Sadovnikovia belemnoides9. В первоописании садовниковия отнесена к семейству Pleuromeiaceae. Первоначально весь материал, положенный в основу описания данного растения, ограничивался единственным спорофиллом, а также двумя фрагментами коры, предположительно принадлежавшими тому же материнскому растению, а также несколькими филлоидами из тех же отложений [8].
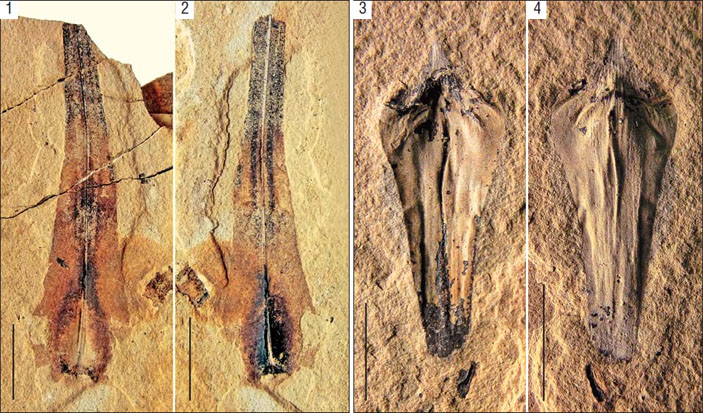
Ископаемые остатки садовниковии Sadovnikovia belemnoides из нижнепермских (кунгурский ярус) отложений Приуралья: 1, 2 — голотип, 3 и 4 — отпечаток и противоотпечаток спорофилла из апикальной части побега
Несмотря на то что садовниковию вполне правомерно отнесли к семейству плевромейевых, в ее строении было много черт, позволяющих сближать ее и с семейством изоэтовых, или полушниковых (Isoetaceae). Более того, у садовниковии наблюдалось такое сочетание признаков, что этот род можно в равной степени отнести и к семейству плевромейевых, и к семейству изоэтовых. Садовниковии были относительно небольшими растениями, спорофиллы которых по спирали прикреплялись к относительно короткой центральной оси. В самой нижней части побега могли располагаться стерильные листья — филлоиды.
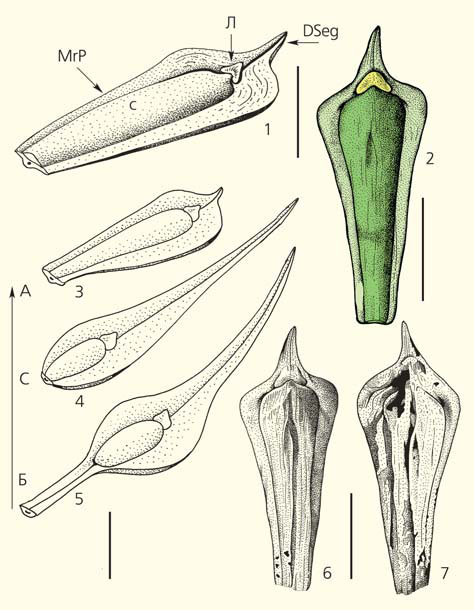
Интерпретация морфологии садовниковии Sadovnikovia belemnoides: 1 — строение спорофилла, вид сбоку (обозначены: DSeg — дистальный сегмент, апекс спорофилла, Л — лигула; MrP — краевая часть пластинки спорофилла, С — спорангий); 2 — реконструкция спорофилла; 3 — спорофилл из апикальной части побега (A — апикальная часть); 4 — спорофилл из средней части побега (С — средняя часть); 5 — спорофилл из базальной части побега (Б — базальная часть), стрелкой отмечено направление к верхушке побега; 6, 7 — прорисовки отпечатка и противоотпечатка спорофилла из апикальной части побега
Таким образом, плевромейевые (считая садовниковию их представителем) появились по меньшей мере за 30 млн лет до момента, когда это событие должно было произойти, согласно представлениям Мэгдефрау. И более того, присутствие в строении садовниковии отчетливых черт строения шильника (изоэтеса) указывало на то, что новый род мог быть общим предком и плевромейевых, и изоэтовых, что в корне подрывало концепцию немецкого палеоботаника. Получилось, что предполагавшаяся филогенетическая последовательность в эволюции гетероспоровых плауновидных нарушалась очень существенно. Растения, которым по схеме Мэгдефрау полагалось произрастать на Земле лишь в мезозое, вполне хорошо чувствовали себя в первой половине пермского периода в Приуралье.
Но на этом сложности в систематике гетероспоровых плауновидных не закончились. Находка садовниковии в Чекарде позволила взглянуть по-новому на некоторые ранее найденные растения. Одним из таких растений был томиостробус. Расскажу о нем поподробнее.
В середине прошлого века из нижнетриасовых отложений Сибири было описано интересное растение, названное Tomiostrobus radiatus. Первоначально его отнесли к араукариевым, но после детального изучения выяснилось, что это спорофиллы гетероспорового плауновидного (подробнее см.: [9]). Таким образом, в филогенетическом ряду от палеозойских гетероспоровых плауновидных к современному шильнику появился еще один элемент.

Реконструкция садовниковии Sadovnikovia belemnoides
Первые материалы по томиостробусу были относительно малочисленными, но новые массовые сборы спорофиллов этого растения позволили предложить реконструкцию томиостробуса [10]. Согласно ей, триасовый томиостробус выглядел гораздо более похожим на современный изоэтес, чем раннемеловая натгорстиана. Эти данные косвенно подтверждаются сведениями и о других триасовых гетероспоровых плауновидных, родственных современному шильнику или даже непосредственно отнесенных к роду Isoetes [11, 12].

Реконструкция садовниковии. Часть спорофиллов удалена, показано продольное сечение побега
Таким образом, общая концепция эволюции гетероспоровых плауновидных поменялась очень существенно. Выяснилось, что уже в раннем триасе существовали настоящие представители семейства изоэтовых, а в раннепермскую эпоху в Приуралье произрастали плауновидные (Sadovnikovia), которых можно рассматривать как переходную форму от семейства плевромейевых к семейству изоэтовых, или форму, находящуюся в основании дивергентного события, ведущего к изоэтовым и плевромейевым.
Теперь нам остается поискать новых представителей семейства плевромейевых из пермских отложений. Некоторые из них нам уже известны. Это роды Viatcheslavia и Signacularia. Но похоже, что есть и более древние представители плевромейевых. Эти находки еще находятся в стадии изучения, которое позволит пролить новый свет на филогенетическую историю гетероспоровых плауновидных.
***
В геологической истории таксонов ранга семейства (и отчасти более высоких) ученые часто имеют дело с уже достаточно хорошо обособленными группами, причем даже в тех случаях, когда их близкое родство совершенно очевидно.
После дивергентного расхождения такие семейства уже прошли значительный эволюционный путь, и они разделены широким и хорошо читающимся таксономическим / морфологическим интервалом. Однако совершенно очевидно, что в условный момент дивергентного события морфологический интервал между сестринскими семействами был незначительным. Вполне можно допустить, что признаки новых семейств проявлялись в разных родах мозаически, в разных сочетаниях, с разной частотой и последовательностью. Более того, очевидно, что в генотипе двух новых семейств (т.е., в контексте вышеизложенного, плевромейевых и изоэтовых) сохранялись гены предкового семейства, которые могли оставаться в латентном состоянии и, будучи рецессивными признаками, никак не проявляться долгое время, но затем вновь так или иначе отразиться в фенотипе.
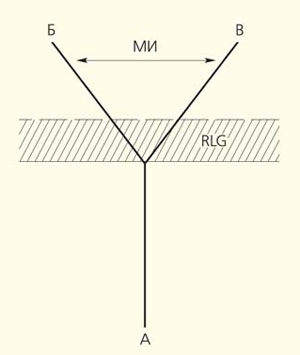
Предполагаемая схема происхождения семейств Pleuromeiaceae (Б) и Isoetaceae (В) от гипотетической предковой группы (A). RLG (recapitulation of latent genes, англ.) — рекапитуляция латентных генов, унаследованных плевромейевыми и изоэтовыми от общего предка. МИ — морфологический интервал
Подобная плохо читаемая в палеонтологической летописи информация может сильно осложнять интерпретацию, особенно при анализе непосредственной окрестности дивергентного события. В таких сложных случаях палеонтологи нередко прибегают к таксономическим приемам, страхующим от ошибок. Создается условная или параллельная «паратаксономия» для упорядочивания морфологически сходных форм, степень естественного родства которых неясна или неочевидна.
В случае с семействами Pleuromeiaceae и Isoetaceae эта проблема стоит не столь остро, хотя самые древние предковые формы обоих семейств очень сходны друг с другом, а на фрагментарных остатках подчас и неразличимы.
Какие любопытные палеоботанические выводы можно сделать из всего вышеизложенного? Прежде всего, благодаря новым находкам удалось установить, что семейство изоэтовых появилось не позднее раннетриасовой эпохи. Вероятно, изоэтовые существовали и значительно раньше, в середине пермского периода. Именно в то время в Приуралье произрастал возможный общий предок изоэтовых и плевромейевых — садовниковия. Но пока нельзя считать позднепалеозойскую историю гетероспоровых плауновидных полностью расшифрованной. Возможно, новые находки помогут прочитать неизвестные нам страницы геологической летописи эволюции этих интересных растений.
Работа выполнена в рамках темы госзадания № 0135-2019-0044 «Фитостратиграфия, палеофлористика, кризисные события кайнозоя, мезозоя и палеозоя различных регионов Евразии, палеоклиматические, палеофитоценотические и палеофитогеографические реконструкции» Геологического института РАН, а также за счет средств субсидии, выделенной в рамках государственной поддержки Казанского (Приволжского) федерального университета в целях повышения его конкурентоспособности среди ведущих мировых научно-образовательных центров.
Литература
1. Наугольных С. В. Первая находка лианоподобного птеридосперма в нижнем карбоне Урала // Природа. 2018; 3: 87–88.
2. Naugolnykh S. V., Orlova O. A. Moscvostrobus — a new genus of Carboniferous lycopods from the Moscow region (Russia) // The Palaeobotanist. 2006; 55: 1–14.
3. Mägdefrau K. Über Nathorstiana , eine Isoëtaceae aus dem Neokom von Quedlinburg am Harz // Beihefte zum botanischen Centralblatt. 1932; 49(II): 706–718.
4. Mägdefrau K. Paläobiologie der Pflanzen. Jena, 1956.
5. Тахтаджян А. Л. Высшие растения. I: От псилофитовых до хвойных. М.; Л., 1956.
6. Пономарева Г. Ю., Новокшонов В. Г., Наугольных С. В. Чекарда — местонахождение пермских ископаемых растений и насекомых. Пермь, 1998.
7. Жужгова Л. В., Пономарева Г. Ю., Аристов Д. С., Наугольных С. В. Чекарда — местонахождение пермских ископаемых насекомых и растений: Монография по геологии, палеоэнтомологии и палеоботанике Чекарды. Пермь, 2015.
8. Naugolnykh S. V. A new lepidophyte from the Kungurian of the Central Urals // Paleontological Journal. 1994; 28(2): 168–175.
9. Садовников Г. Н. Морфология, систематика и распространение рода Tomiostrobus // Палеонтологический журнал. 1982; 1: 100–109.
10. Naugolnykh S. V. Sporophyll morphology and reconstruction of the heterosporous lycopod Tomiostrobus radiatus Neuburg emend. from the Lower Triassic of Siberia (Russia) // Palaeobotanist. 2012; 61: 387–405.
11. Retallack G. J. Earliest Triassic origin of Isoetes and quillwort evolutionary radiation // Journal of Paleontology. 1997; 71(3): 500–521.
12. Наугольных С. В. Могучева Н. К. Новый представитель плауновидных из нижнего триаса Тунгусской синеклизы (Сибирская платформа) // Новости палеонтологии и стратиграфии: Приложение к журналу «Геология и геофизика». 2006; 8: 81–93.
1 Филлоид — стерильный лист плауновидного или мохообразного, образованный выростами поверхностных тканей.
2 Плауноводные // Биологический энциклопедический словарь / Гл. ред. М. С. Гиляров. М., 1989. С. 478.
3 Терминальный стробил — компактное собрание (стробил или шишка) спорофиллов, расположенное на конце побега.
4 Апикальная меристема — зона роста, расположенная на верхушке побега.
5 Дистальное положение — расположение части какого-либо органа в отдалении от основания этого органа; дословно — «дальнее» положение.
6 Мейен С. В. Из истории растительных династий. М., 1971; Он же. Следы трав индейских. М., 1981.
7 Пролиферация — разрастание тканей в организме; в морфогенезе побега — прорастание побега выше стробила или фертильной зоны.
8 Фертильная зона — собрание спорофиллов, выше которого располагается нормальный вегетативный побег с листьями.
9 Родовое название было дано по имени известного российского палеоботаника Г. Н. Садовникова, посвятившего ряд своих работ систематике и морфологии гетероспоровых плауновидных. Видовой эпитет образован от древнегреческого слова belemnon — ‘стрела’, поскольку морфология спорофилла напоминала очертания наконечника античной стрелы.














Selaginella — современное гетероспоровое плауновидное. Здесь и далее фото автора