Изотопные маркеры жизни древних людей
Мария Добровольская, Ирина Решетова
«Природа» №1, 2019
 Мария Всеволодовна Добровольская — доктор исторических наук, ведущий научный сотрудник отдела теории и методики Института археологии РАН. Область научных интересов — палеоантропологический источник в археологических исследования, экология человека, палеодиетологические реконструкции. |
 Ирина Константиновна Решетова — кандидат исторических наук, научный сотрудник того же отдела института. Специалист в области биоархеологических и палеоэкологических реконструкций, палеоантропологии, одонтологии, палеодемографии. |
Палеоэкологические исследования — одно из актуальных направлений в современной археологии. Получить надежные факты, характеризующие природную и антропогенную среду, в которой проходила жизнь людей, создать обоснованную и понятную широкому кругу специалистов картину окружения человека прошлого возможно только на базе тесного сотрудничества между представителями различных наук.
С середины XX в. в исторической науке стали популярны новые аспекты исследований, объединенные под единым названием «история повседневности». Это связано с общим кризисом исторической науки, а также с приходом новых поколений исследователей. Становится ясно, что исторический процесс далеко не полностью отражен в официальных исторических событиях, ярких социальных, политических, технологических и прочих достижениях, запечатленных в памяти народа, благодаря их очевидной значимости и экстраординарности.
Поле изучения истории повседневности — условия и качество жизни, специфика каждодневного труда, традиции питания, лечения и прочие сферы быта, составляющие его обычное фактическое содержание. Отметим, что археология как историческая дисциплина (связанная с изучением, в частности, вещественных источников) описывает обыденные события прошлого, в той или иной мере всегда воссоздает страницы повседневной жизни людей, отделенной от нас многими столетиями и тысячелетиями.
Одна из важнейших сторон жизни — еда. То, что ели наши предшественники, как, когда и по каким поводам, и как это было обусловлено их культурой, а также природными ресурсами, типом хозяйства — поле обширного и разностороннего исследования в современной археологии. Один из популярных подходов, получивший развитие в археологии в последние десятилетия прошлого века, связан с привлечением знаний из области фундаментальной биологии, на базе которых возник метод, позволяющий реконструировать индивидуальные усредненные пищевые рационы. Этот подход — один из ярких примеров конвергенции научных направлений, на базе которых возникают новые знания. Конечно, лучше всего было бы обзавестись чудесным горшочком из сказки Андерсена, чтобы, держа руку над подымающимся из него паром и слушая перезвон колокольчиков, определять: где и что у кого было на ужин. Однако за неимением последнего мы обратимся к данным биохимии и экологии.
От изотопных соотношений к пищевым пристрастиям
В основу метода положены сведения об изотопном составе элементов, которые входят в структуру белков, углеводов и жиров. Остановимся на изотопном составе двух элементов: углерода и азота. Согласно общеизвестным представлениям о пищевых цепях, весь углерод и азот из органических соединений, попадающий в пищеварительную систему, мы получаем из растений (продуцентов) и животных (консументов различных уровней). Изотопный состав этих важнейших химических элементов различен во всех звеньях трофических цепей. Причин тому множество, но наиболее сильные трансформации изотопный состав претерпевает в процессе фотосинтеза. Как известно, наиболее распространены два механизма фотосинтеза. Один (цикл Кальвина) формирует молекулу с тремя атомами углерода (С3), и по этому типу фотосинтеза «работает» большинство травянистых, кустарниковых и древесных растений. Другой (цикл Хэтча — Слэка) формирует молекулу с четырьмя атомами углерода (С4), и он характерен для некоторых растений аридных районов (сахарный тростник, кукуруза, просо). В результате фотосинтеза изотопный состав углерода, получаемого растениями из воздуха или воды, изменяется в сторону уменьшения доли тяжелых атомов.
Так как тяжелые изотопы азота и углерода содержатся в ничтожных количествах, принято оценивать изотопное соотношение в величинах дельта (δ), которые измеряются в промилле (‰) и высчитываются по следующим формулам:
δ13C = [[(13C/12Cобразца)/(13C/12Cстандарт)] − 1] × 1000;
δ15N =[[(15N/14Nобразца)/(15N/14Nстандарт)] − 1] × 1000.
В качестве стандарта для азота используется атмосферный азот, а для углерода — Pee Dee Belemnite (PDB) из морских окаменелостей мелового периода Belemnitella americana из формации Пии Дии в Южной Каролине. Эти образцы имеют аномально высокое отношение 13C/12C (0,0112372) и приняты в качестве эталона нулевого значения δ13C. Использование этого стандарта приводит к отрицательным значениям δ13C для обычных материалов.
Для каждого из этих типов фотосинтеза существуют свои границы изменчивости δ13С: для С3 — от −35 до −22‰, а для С4 — от −17 до −9‰ [1]. Отправная точка для наземных экосистем — величина δ углекислого газа воздуха. Величина δ13C воздушного углерода составляет около −7‰. Морские экосистемы имеют свою специфику. Значение δ13C из морских биокарбонатов составляет 0‰. Поэтому, в зависимости от насыщенности морской воды древним биокарбонатным углеродом, величины δ13C фитопланктона будут значительно варьировать (от −31 до −7‰). В среднем показатели по фитопланктону составляют −18‰ [2]. Величина трофического «шага» δ13C при переходе от одного трофического уровня к другому варьирует от 0,8 до 5‰. Согласно предположениям С. Амброза и Л. Норра [3], эта величина зависит от размеров тела организма-консумента, его метаболизма и структуры питания. Для человека, как правило, величина трофического шага приближается к максимальной величине — 5‰.
Путь азота в пищеварительный тракт такой же, как и углерода: через звенья трофической цепочки — от растений и животных. В растения азот попадает из почвы (как правило), но есть и азотфиксирующие растения, способные поглощать этот элемент из воздуха. Изотопный состав азота обычных растений зависит от особенности почвы, а состав азота в тканях азотфиксирующих растений приближен к изотопному составу азота воздуха (около 0‰). Наземные и морские экосистемы также различаются по изотопному составу азота. Величина δ15N азота океанических систем много выше, так как доля тяжелых изотопов там больше. Протеин морского происхождения характеризуются значениями δ15N от 14 до 20‰ [4], в то время как изотопное соотношение азота мышечной ткани травоядных наземных животных может падать до 3–4‰. Величина трофического шага δ15N в трофических цепочках значительно варьирует и в среднем составляет 3‰. Существенные изменения изотопного состава азота происходят при переходе с одного трофического уровня на другой в аридных экосистемах (до 5–6‰). В гумидных экосистемах величина такого изменения не превышает 3‰.
Таким образом, в трофических цепях различных экосистем происходят процессы обогащения и обеднения тяжелыми изотопами азота и углерода. Интересно отметить, что еще в 1926 г. в своей работе «Изотопы и живое вещество» В. И. Вернадский писал: «Живые организмы способны избирать определенные изотопы из их смесей, каковыми являются многие элементы окружающей нас среды» [5, с. 85]. В процессе фотосинтеза происходит фракционирование, в результате которого доля тяжелых изотопов углерода убывает в большой или меньшей степени, в зависимости от типа фотосинтеза.
Итак, знания о значительных изменениях изотопного состава азота и углерода в процессе фотосинтеза и фракционирования в звеньях трофический цепей позволяют нам реконструировать основные пищевые ресурсы человека или животного, если известен экологический контекст его существования. На этом и основаны биоархеологические палеодиетологические реконструкции.
Как выстраиваются конкретные реконструкции пищевых предпочтений прошлого? Обратимся к примерам.
Лесные жители
Смешанные, южнотаежные и таежные леса европейской части России в эпоху позднего каменного века были активно освоены человеком. Наиболее притягательными местами для поселений были побережья озер, особенно в устьях или истоках рек [6]. Льяловская неолитическая культура (4–3-е тысячелетия до н. э.) известна прежде всего массивными керамическими сосудами с заостренным дном, покрытыми ямочно-гребенчатым орнаментом [7]. Ей на смену приходит волосовская культура (3–2-е тысячелетия до н. э.), в среде которой уже формируются ранние формы металлургии, — таким образом, она завершает бытование культур каменного века. Зачастую носители традиций обеих этих культур выбирали для поселений (равно как и для погребений) одни и те же места, в результате чего формировались многослойные археологические памятники, содержащие информацию о многотысячелетней истории.
Один из таких памятников — могильник Сахтыш 2а, расположенный в Тейковском р-не Ивановской обл. [8]. На его площади были обнаружены многочисленные погребения волосовской и льяловской культур. Сохранность скелетных останков позволила нам провести определения изотопных соотношений углерода и азота в коллагене костной ткани людей.
Прежде всего обратим внимание на то, что локализация показателей для индивидов двух разных культур не совпадает. Величина δ15N, определенная для образцов из погребений льяловской культуры, варьирует от 12 до 14,6‰, а для образцов представителей волосовской культуры — от 11 до 13‰ [9]. Границы значения δ13C для образцов из скелетных тканей представителей волосовской культуры составляют диапазон от −24 до −20,5‰, в то время как изотопное соотношение углерода для льяловцев всегда выше −21,5‰, а один образец (погребение № 29) характеризуется величиной −17,6‰! Это свидетельствует о различиях в пищевых пристрастиях представителей этих двух археологических культур. У всех индивидов величины δ15N умеренные и высокие, что однозначно указывает на заметное место белковой пищи в обыденном рационе этих людей. Единственный образец местной древней фауны (вероятно, бобр) характеризуется гораздо более низким значением δ15N (6,8‰).
Графическая реконструкция (выполнена Г. В. Лебединской) облика женщины из погребения 22 могильника Сахтыш 2а, льяловская археологическая культура
Сравним с характеристиками других неолитических людей. Величина δ13C для образцов костной ткани из неолитических погребений, оставленных обитателями побережий озер с территории современной Латвии (памятник Звейниеки), варьирует от −24 до −21,5‰, а δ15N — от 12 до 13,5‰; мезолитические рыболовы, жившие на берегах Днепра (могильник Васильевка 3), характеризуются δ13C около −22‰ и δ15N около 14‰.
Все это позволяет сделать вывод, что население, оставившее погребения волосовской культуры, судя по всему, активно занималось рыбной ловлей, причем вряд ли это была рыба крупных размеров. Реконструкция питания представителей льяловской культуры вызывает много вопросов. Можно предполагать, что охота, а не рыбный промысел была ведущей деятельностью в системе их жизнеобеспечения. Индивид, характеризующийся величиной дельта углерода −17,6‰, вероятнее всего, был связан с другими, более аридными экосистемами, так как высокие значения δ13C однозначно указывают на роль растений типа фотосинтеза С4 в трофической цепочке, в которую оказался данный человек включен. Либо его обыденный рацион был представлен пищей специфического изотопного состава углерода. Оказали влияние на выбор пищевых ресурсов изменения климата суббореального периода? Была ли традиция хозяйственного уклада фактором, корректирующим пищевые предпочтения? Существовали ли у льяловцев контакты с более южными открытыми пространствами? Все эти вопросы требуют дальнейших исследований. Полученные результаты убедительно демонстрируют, что условный термин «охотники-рыболовы-собиратели» подразумевает множество разнообразных стратегий хозяйствования и питания, без изучения которых наши знания об образе жизни первобытных неолитических жителей лесных пространств будут во многом формальны.
Пространственное расположение памятников льяловской и волосовской культур, изученное ранее, показало, что для последней характерны более крупные и стационарные поселения в местах, наиболее подходящих для эффективного массового вылова рыбы, а поселения более ранней неолитической культуры такой специализации не имеют. Таким образом, археологические сведения и данные изотопных исследований подтверждают друг друга. Но от каждого метода мы ждем новых возможностей, которые могут не только подтверждать или опровергать уже высказанные гипотезы, но и формулировать новые вопросы. Изотопные исследования позволяют обсуждать индивидуальные особенности усредненных пищевых рационов, что было невозможно до появления подобных методов. Характеризовать индивидуальность очень важно для понимания социальных аспектов жизни древних коллективов. На примере людей из погребений могильника Сахтыш 2а мы видим, что индивидуальные значения δ13C могут различаться почти на 4‰, т. е. на экологически значимую величину. Таким образом, возможно, существовали традиции или обстоятельства, которые способствовали формированию индивидуальных особенностей питания в первобытном социуме.

Изотопные показатели — значения δ13С и δ15N, определенные в коллагене костной ткани неолитических людей из могильника Сахтыш 2а. Красным обозначены показатели представителей льяловской культуры, а синим — волосовской культуры
Наши представления о социальной жизни охотников-рыболовов-собирателей крайне ограниченны, однако все больше и больше появляется указаний, которые свидетельствуют о сложности их хозяйственных и социальных укладов. Это, безусловно, отдаляет нас от мнения о первобытных обществах каменного века как о «примитивных» и «простых». Возможно также, что видимая нами индивидуальная изменчивость обусловлена широким разнообразием пищевых источников, используемых древними. Можно предположить, что понятие «охотники-собиратели» объединяет во многом различающиеся системы жизнеобеспечения, а члены этих социумов существовали вне жестких стандартов образа жизни. Охотники-рыболовы-собиратели позднего каменного века тысячелетиями жили в окружении лесов и водоемов. Они приспосабливались к богатым природным ресурсам, а не трансформировали и унифицировали их коренным образом через сельскохозяйственную деятельность. И при этом численность людей неуклонно росла на протяжении всего неолита. Мы можем рассматривать это как пример оригинального пути развития, на котором не могла возникнуть идея о «победе над природой», так как она была бы самоубийственной.

Слева: графическая реконструкция (выполнена Г. В. Лебединской) облика мужчины из погребения 40 могильника Сахтыш 2а, льяловская археологическая культура. Справа: графическая реконструкция (выполнена Г. В. Лебединской) облика мужчины из погребения 39 могильника Сахтыш 2а, волосовская археологическая культура
Древнейшие пастушеские племена на Среднем Дону
Эпоха бронзы в степных и лесостепных ландшафтах Евразии (условно, 4–2-е тысячелетия до н. э.) была временем масштабных миграций и распространений новых технологий: металлургии, скотоводства, земледелия. Так, палеогенетические исследования последних лет наглядно продемонстрировали значительное изменение генетического состава европейского населения в эпоху бронзы. Именно в то время закладывался современный генетический фундамент Европы. Одна из крупных культурно-исторических общностей эпохи бронзы (3–2-е тысячелетия до н. э.) — катакомбная. Она ассоциируется с продвижением культурных и миграционных импульсов южного происхождения в степные и лесостепные регионы Восточной Европы. На Среднем Дону эта культура оставила многочисленные курганные могильники и следы стационарных поселений [10]. Мы провели определения значений δ15N и δ13С в коллагене костной ткани людей из погребений среднедонской катакомбной культуры (Воронежская обл.).

Значения δ13С и δ15N в коллагене костной ткани людей и мелкого рогатого скота (одно животное) из курганных погребений среднедонской катакомбной культуры (могильники Власовский 2, Репная Балка, Колбино 1). Малый красный овал объединяет показатели основной группы индивидов, а большой синий — включает область значений всех индивидов
Преобладающее число индивидов характеризуются устойчивыми показателями δ13С в пределах от −19 до −20‰ и δ15N от 10,5 до 12‰. Величины δ13С соответствуют флоре типа фотосинтеза С3, но в уже другом ее варианте — лесостепном. Вариации δ15N однозначно указывают на преобладание белкового компонента в их рационе. В качестве единственного «репера», характеризующего хозяйственные связи, мы проанализировали кость от скелета взрослой овцы или козы из погребения. Значение δ13С в образце было аналогично величинам, определенным для людей, а вот значение δ15N было низким, что полностью соответствует представлению об изотопном соотношении азота в коллагене кости травоядного животного.
Основываясь на палеодиетологической реконструкции, можно сделать вывод, что подавляющее большинство членов сообщества среднедонской катакомбной культуры вело очень консолидированный образ жизни, используя небольшой спектр пищевых источников. Представления об их пастушеской деятельности позволяют конкретизировать: их пищей было мясо и молоко домашних животных.
Для двух мужчин мы провели сопоставление изотопного состава коллагена корня зуба и фрагмента компактной части трубчатой кости. Дело в том, что в скелете можно различать консервативные и реактивные типы тканей. Первые формируют свой состав по мере роста, а затем мало его изменяют. Вторые постоянно перестраиваются и поэтому отражают в своем составе события жизни последних нескольких лет. К первым могут быть отнесены твердые ткани корней зубов человека, а ко вторым — компактная часть трубчатых костей. Корни зубов (вторых моляров), которые были изучены, формируются в детстве и отрочестве (примерно от 8 до 13 лет). В нашем распоряжении были две пары образцов зуб / кость. Первая пара относилась к юноше 15–19 лет из кургана памятника Репная Балка, величины δ13С и δ15N в кости и корне зуба практически совпали. Вторая пара была получена из скелета мужчины того же курганного некрополя, но его возраст 40–49 лет. И тут мы видим незначительное снижение показателей по обоим элементам. Но уловить смену модели питания на протяжении нескольких лет жизни двух человек нам не удалось.

Графическая реконструкция (выполнена И. К. Решетовой) облика мужчины из погребения 83 могильника Ксизово 19, катакомбная культурно-историческая общность, Верхний Дон
Следует отметить, что на фоне монотонных показателей выделяются индивиды, которые характеризуются повышенными значениями δ13С, что позволяет включать их в трофические цепи с участием растений типа фотосинтеза С4. Почему это важно? Для Евразии эпохи бронзы просо — единственное культурное растение, которое может быть отнесено в группе типа фотосинтеза С4. Вопрос о традиции употребления проса в пищу интересен как в связи с реконструкцией хозяйства катакомбных племен на Среднем Дону, так и в связи с возможным «восточным следом» и, предположительно, существованием своего рода «Изотопного Просяного пути» из Китая в Европу через всю Евразию [11]. Значения δ13С, полученные для двух людей из курганного могильника Колбино 1 и одного человека из курганного могильника Богучарский 2 соответствуют величинам, определенным для памятников позднебронзовой карасукской культуры Минусинской котловины [12].
Существуют указания на независимый центр возделывания проса в раннем бронзовом веке в Северной Италии, а затем на постепенное распространение в Восточную Европу зоны культивирования этого растения [13]. В любом случае вызывает большой интерес спорадическое появление отдельных людей с высокими показателями δ13С. Так, на примере трех человек из одного кургана могильника Колбино 1 мы фиксируем двух мужчин с высоким показателем δ13С и одного с умеренным, типичным для большинства погребенных. При этом значение δ15N для всех трех индивидов примерно одинаково.
Палеодиетологическая реконструкция позволяет нам сформулировать гипотезу о достаточно стабильном существовании пастушеских племен на территории Среднего Дона. Это подтверждается и археологическими данными, указывающими на существование сезонных поселений, которые позволили в условиях пересеченного рельефа наиболее эффективно использовать пастбищные возможности на относительно небольшой территории. Спорадическое появление отдельных индивидов с высокими значениями δ13С пока требует дальнейших исследований. Возможно, что отдельные проявления употребления проса — следы связей внутри этой обширной области.
Номады раннего железного века
В середине 1-го тысячелетия до н. э. на земли Среднего Дона приходят номады (кочевники) — носители традиций среднедонской культуры скифского времени — и возводят свои курганы. Нам показалось интересным сравнить своеобразие изотопного состава коллагена костной ткани двух совершенно несхожих обществ, живших на одних и тех же землях, но отделенных друг от друга более чем тысячелетием. Люди, хоронившие своих соплеменников под курганными насыпями в раннем железном веке, принадлежали к кругу скифских культур, о чем свидетельствует богатое сопровождение оружием, украшениями из золота, которым снабжались тела при погребальных церемониях [14].

В серию из 29 образцов костной ткани, полученных от скелетных останков, что погребены в могильнике Колбино-Терновое, были включены взрослые обоих полов и дети [15]. Диапазон изменчивости δ13С составил от −20,5 до −13,8‰. Хотя внутри этих широких границ есть самые разные варианты δ13С, выявить какие-либо группировки нам не удалось. Столь широкие пределы изменчивости однозначно свидетельствует о существовании значительных индивидуальных вариаций в употреблении растительной пищи. Именно индивидуальных: если бы основу питания составляла смешанная растительная пища (включающая растения типов фотосинтеза С3 и С4), то величины δ13С были бы примерно сходны и находились в зоне промежуточных значений. Между тем мы имеем именно множество вариантов растительной основы рационов: от только С3 до только С4. Значение δ15N варьирует от 10,7 до 12,7‰, уверенно находясь в зоне «преимущественно белковой пищи». Полученная картина не оригинальна. Так, для скифских погребений с территории Тувы (памятник Аймыр-Лыг) были также показаны высокие значения δ13С в диапазоне изменчивости около 4,5‰ и высокие значения δ15N [16]. Можно заключить, что вне зависимости от локализации той или иной группы культурные традиции питания приводят к сходным картинам внутригрупповой изменчивости δ13С и δ15N.
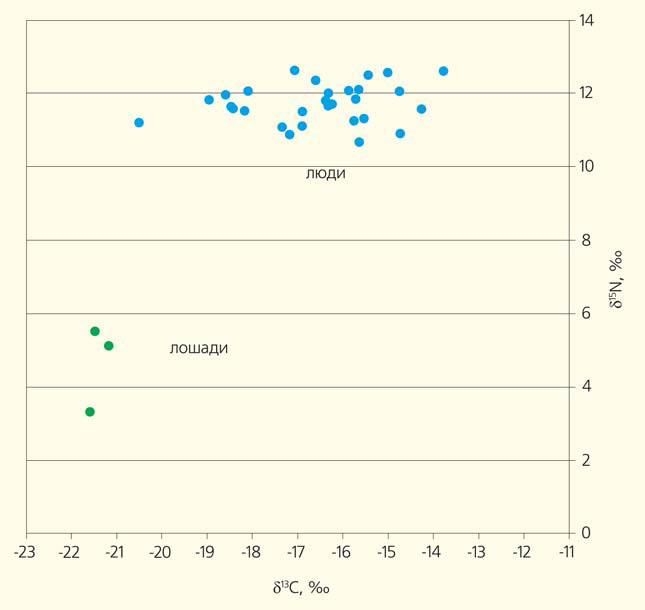
Значения δ15N и δ13С, определенные для образцов коллагена людей и лошадей (IV в. до н. э.), курганные погребения скифской культуры на Среднем Дону
Культура питания
На примере двух столь разных обществ — из эпохи бронзы и из раннего железного века, хозяйство которых было в основе своей связано со скотоводством, мы видим совершенно различные социальные тенденции, формирующие традиции питания. Если для ранних скотоводов можно предположить консолидированный образ жизни социума, использование (за редким исключением) определенных пищевых ресурсов, то для общества раннего железного века мы наблюдаем большое индивидуальное разнообразие изотопных показателей.
Попытки реконструировать особенности питания в обществах прошлого базируются, с одной стороны, на сугубо экологических закономерностях формирования локальных трофических сетей, а с другой — на представлении о том, что питание — это культурный феномен. В этом отношении изотопные методы изучения питания носят характер междисциплинарных, так как позволяют получать данные методами естественных наук и использовать их для изучения культурного своеобразия, социальной структуры обществ, форм хозяйства и образа жизни.
Наши исследования требуют и совершенствования методик, и привлечения большего объема материалов, и развития теоретической базы. Однако уже сделанные шаги свидетельствуют о том, что у археологов появляется еще одна возможность изучать обыденную жизнь людей, пытаясь реконструировать не только усредненные, «типичные», характеристики, но и частности, как бы возвращая из прошлого многообразные и неповторимые черты индивидуальных жизней.
Литература
1. Tykot R. H. Stable isotopes and diet: You are what you eat. Proceed. of the Intern. School of Physics «Enrico Fermi» Course. Martini M., Milazzo M., Piacentini M. (eds.). Amsterdam, 2004: 433–444.
2. Merve N. van der. Carbon isotopes, photosynthesis, and archaeology // American Scientist. 1982; 70: 596–606.
3. Ambrose S., Norr L. Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen carbonate. Prehistoric human bone — Archaeology at the Molecular level. Lambert J. B., Grupe G. (eds.). Berlin, 1993.
4. Shoeninger M. J., DeNiro M. J. Nirtogen and carbon isotopic composition of bone collagen from marine and terrestrial animals // Geochemica et Cosmochimica Acta. 1984; 48: 625–639.
5. Вернадский В. И. Изотопы и живое вещество // Вернадский В. И. Биогеохимические очерки. М.; Л., 1940; 84–88.
6. Сидоров В. В., Энговатова А. В. Основания палеоэкономических реконструкций присваивающего хозяйства. Тверской археологический сборник. Вып. 2. Тверь, 1996; 15–26.
7. Энговатова А. В. Хронология эпохи неолита Волго-Окского междуречья. Тверской археологический сборник. Вып. 3. Тверь, 1998; 238–246.
8. Крайнов Д. А., Костылева Е. Л., Уткин А. В. Могильник и «святилище» на стоянке Сахтыш 2а. Российская археология. 1994; 2: 118–130.
9. Энговатова А. В., Добровольская М. В., Костылева Е. Л. Изотопные характеристики индивидов из погребений неолитического могильника Сахтыш 2а: данные к реконструкции питания. Археология Подмосковья. Вып. 11. М., 2015; 138–146.
10. Гак Е. И. Поселение Рыкань 3 в контексте памятников катакомбной общности восточневропейской лесостепи. Краткие сообщения Института археологии. 2013; 230: 141–161
11. Tingting W., Dong W. Chang X. et al. Thianshanbeilu and the Isotopic Millet Road: reviewing of late Neolithic / Bronze Age radiation of human millet consumption from North China to Europe / National Science Review. 2017; 00: 1–16. DOI: 10.1093/nsr/nwx015.
12. Святко С. В. Анализ стабильных изотопов: основа метода и обзор исследований в Сибири и Евразийской степи. Археология, этнография и антропология Евразии. 2016; 44(2): 47–55.
13. Tafuri M. A., Craig O. E., Canci A. Stable isotope evidence for the consumption of millet and other plants in Bronze Age Italy // American Journal of Physical Anthropology. 2009; 139(32): 146–153.
14. Гуляев В. И. На восточных рубежах Скифии (древности донских скифов). М., 2010.
15. Добровольская М. В., Решетова И. К., Чагаров О. С. Стабильные изотопы в реконструкции традиций питания населения Среднего Дона в эпоху бронзы и раннем железном веке. V (XXI) Всероссийский археологический съезд. Барнаул, 2017; 321–322.
16. Eileen M. Murphy E. M. Schulting R. et al. Iron Age pastoral nomadism and agriculture in the eastern Eurasian steppe: implications from dental palaeopathology and stable carbon and nitrogen isotopes // Journal of Archaeological Science. 2014; 40: 2547–2560.













Графическая реконструкция (выполнена Г. В. Лебединской) облика женщины из погребения 20 могильника Сахтыш 2а, льяловская археологическая культура