Пионовая тайга
Ольга Смирнова, Алексей Алейников, Дарья Луговая, Николай Смирнов
«Природа» №2, 2014
 Ольга Всеволодовна Смирнова — доктор биологических наук, профессор, заведующая лабораторией структурно-функциональной организации и устойчивости лесных экосистем Центра по проблемам экологии и продуктивности лесов РАН. Область научных интересов — популяционная экология, палеоэкология, теория сукцессии и климакса, история природопользования, рациональное природопользование. |
 Алексей Александрович Алейников — кандидат биологических наук, старший научный сотрудник той же лаборатории. Область научных интересов — структурно-функциональная организация бореальных лесов, история природопользования Урала, средопреобразующая деятельность ключевых видов растений и животных, устойчивое лесопользование. |
 Дарья Леонидовна Луговая — кандидат биологических наук, научный сотрудник той же лаборатории. Область научных интересов — природные и антропогенные факторы формирования растительного покрова, потенциальная растительность. |
 Николай Сергеевич Смирнов — младший научный сотрудник отдела мониторинга парниковых газов в природных и антропогенно-нарушенных экосистемах Института глобального климата и экологии Росгидромета и РАН. Область научных интересов — структура, динамика и взаимосвязь со средой бореальных лесов на разных этапах сукцессии, мониторинг парниковых газов от наземных экосистем. |

Таежные леса в настоящее время привлекают пристальное внимание разных специалистов. Мы знаем, что эти леса, расположенные на огромных малодоступных территориях Северного полушария, — ценный экономический ресурс. И в то же время роль их как лесных экосистем на глобальном уровне становится все серьезнее. Связано это со стремительным истреблением тропических и субтропических лесов. Все в большей степени именно северные леса отвечают за биотическую оптимизацию климата и гидрологического режима, за регулирование биогеохимических циклов элементов, поддержание биологического разнообразия и др. [1–3]. Поэтому как никогда прежде необходимо понимать и оценивать экосистемные функции бореальных (северных) таежных лесов, биологическое разнообразие, исследовать историю их развития. Этим вопросам и посвящена наша статья.
Зеленомошная тайга
В результате долгого антропогенного воздействия — рубок, лесного выпаса, пожаров — на огромных пространствах лесов сформировались малопродуктивные растительные сообщества с небогатым составом. В наше время именно такие леса считаются природными. А. И. Толмачев, например, пишет: «Естественность таежной зоны очевидна для каждого. Во внетропических широтах Северного полушария тайга является самым распространенным типом растительности» [4, с. 8]. «Типичной темнохвойной тайге свойствен не только ограниченный набор деревьев-лесообразователей, но и общая бедность состава флоры». Толмачев говорит и о скудости растительного покрова: «...растения, живущие под пологом темнохвойного леса, образуют в своей совокупности лишь ничтожную массу...» [4, с. 11].

Типичная темнохвойная тайга распространена на Русской равнине, на Урале и в Западной Сибири. Такая тайга представляет собой зеленомошно-кустарничковые и зеленомошно-мелкотравные леса — еловые или пихтово-еловые с примесью кедра сибирского (Pinus sibirica). Древесный полог обычно образован деревьями одного поколения, подрост малочислен и слаб, видовой состав трав и кустарничков беден, эколого-ценотическое разнообразие (количество видов, объединенных отношением к факторам среды и типом растительного сообщества) их минимально.
Моховой же покров развит очень сильно. Это зеленые бореальные мхи, отсюда и название лесов — зеленомошные. Мхи образуют мощный слой медленно разлагающейся лесной подстилки, бедной минеральным питанием. Она очень быстро намокает и высыхает. Такой переменный режим увлажнения неблагоприятен для подроста деревьев, многих трав и кустарничков. Кроме того, сухой моховой покров способствует широкому распространению пожаров. Таким образом, условия для смены поколений здесь очень непростые.
Взрослые деревья невелики, корневая система развита слабо, большинство из них поражено грибными болезнями. Старые деревья, погибая, просто обламываются у основания. При этом не происходит выворачивания кома земли и образования бугров и западин, как бывает в других лесах. Постепенно валежины (упавшие мертвые деревья), погрузившиеся в моховой ковер, и сами покрываются зелеными мхами. На их ковре поселяются кустарнички: брусника (Vaccinium vitisidaea), черника (V. myrtillus) и мелкие травы — кислица (Oxalis acetosella), седмичник (Trientalis europaea), майник (Maianthemum bifolium) [5, 6]. Необходимую влагу эти растения получают из разлагающихся валежин, которые, в отличие от мохового ковра, длительно ее сохраняют.
Из деревьев же на валежинах обычно возобновляется только ель — обыкновенная (Picea abies) и сибирская (P. obovata). Лиственные деревья не приживаются ни на валеже, ни в моховом покрове. При такой однородности древесного состава (с господством ели) лесное сообщество часто поражено грибными инфекциями.

Возобновление ели по валежу. Национальный парк «Паанаярви». Северная Карелия

Подзол иллювиально-железистый в чернично-зеленомошном лесу. Фото А. Д. Бовкунова
Зеленомошные леса сформировались под воздействием человека — в результате сплошных или выборочных рубок и пожаров. Их приметы — разрушающиеся пни и подпалины на старых соснах, в том числе и на кедрах. О былых пожарах свидетельствует и состав почвы. Здесь господствуют подзолы — иллювиально-железистые и иллювиально-гумусово-железистые (иллювий, от лат. illuvies, — ‘разлив’, ‘намывная грязь’) и подзолистые почвы. Гумусовый горизонт очень мал, а то и вовсе отсутствует. В почве постоянно содержится уголь. Мы встречали его в большинстве мест в виде слоя между подстилкой и минеральным горизонтом. Слой угля иногда достигает нескольких сантиметров, что говорит о неоднократных пожарах — верховых и низовых [7]. Антропогенный характер большинства пожаров в таежных лесах подтверждается множеством наблюдений и документирован в материалах заповедников [3, 8].
Высокотравные таежные леса

Уголь из почвенных разрезов пихто-ельников зеленомошных. Печоро-Илычский заповедник. Фото М. В. Бобровского
Тайга не ограничивается только темнохвойными зеленомошными лесами. В этом убеждают научная работа в заповедниках, изучение архивов и картографических материалов [7–10]. Многолетние исследования выявили значительное разнообразие типов таежных лесов [11]. Особо выделен уникальный тип — высокотравные таежные леса [12]. Хотя изучать их детально стали относительно недавно (около 15 лет назад), леса эти уже описаны на территориях от Северной Карелии до Урала и в некоторых районах Западной Сибири — от северной до южной тайги. Удивительный участок высокотравных таежных лесов мы обнаружили также за пределами лесного пояса в кустарниковых тундрах, в Горно-Хадатинском государственном заказнике Ямало-Ненецкого автономного округа. На западе и в центре Русской равнины эти леса представлены ельниками высокотравными, а на востоке (а также на Урале и в Западной Сибири) — высокотравными пихто-ельниками с кедром.
Высокотравные леса располагаются на участках с самыми разнообразными элементами рельефа и на подстилающих породах различных типов [6, 12]. Описаны в основном те леса, что приурочены к востоку Русской равнины и к Уралу: вероятно, потому, что общая сохранность лесов здесь лучше, чем на других территориях.

Точечный ареал высокотравных темнохвойных лесов
Эти леса из разновозрастных деревьев разительно отличаются от зеленомошной тайги по большинству признаков. Здесь в напочвенном покрове господствуют цветковые растения, а доля зеленых мхов минимальна, и произрастают они только на валежинах. Преобладает ель, или совместно господствуют ель и пихта. Встречаются береза, осина, другие мелколиственные деревья и кедр. Скопления деревьев чередуются с большими полянами, где растут высокие красиво цветущие травы. Одно из самых красивых и широко распространенных растений бореального высокотравья — пион уклоняющийся, или марьин корень (Paeonia anomala). Это растение мы выбрали в качестве маркера высокотравных лесов. Необычный облик леса с господством пионов определил название этой статьи.
Многоярусность высокотравной тайги
Высокотравным темнохвойным лесам свойственна ярко выраженная ярусность напочвенного покрова. Здесь в первом ярусе господствуют растения бореального разнотравья, которые дополнены крупными папоротниками. Высота растений достигает полутора — двух метров. Наиболее обычны: пион уклоняющийся (P. anomala), аконит северный (Aconitum septentrionale), дельфиниум высокий (Delphinium elatum), воронец красноплодный (Actaea erythrocarpa), недоспелка копьелистная (Cacalia hastata), купальница европейская (Trollius europaeus), скерда сибирская (Crepis sibirica), василистник малый (Thalictrum minus), щитовник австрийский (Dryopteris dilatata), орлячок сибирский (Diplazium sibiricum).
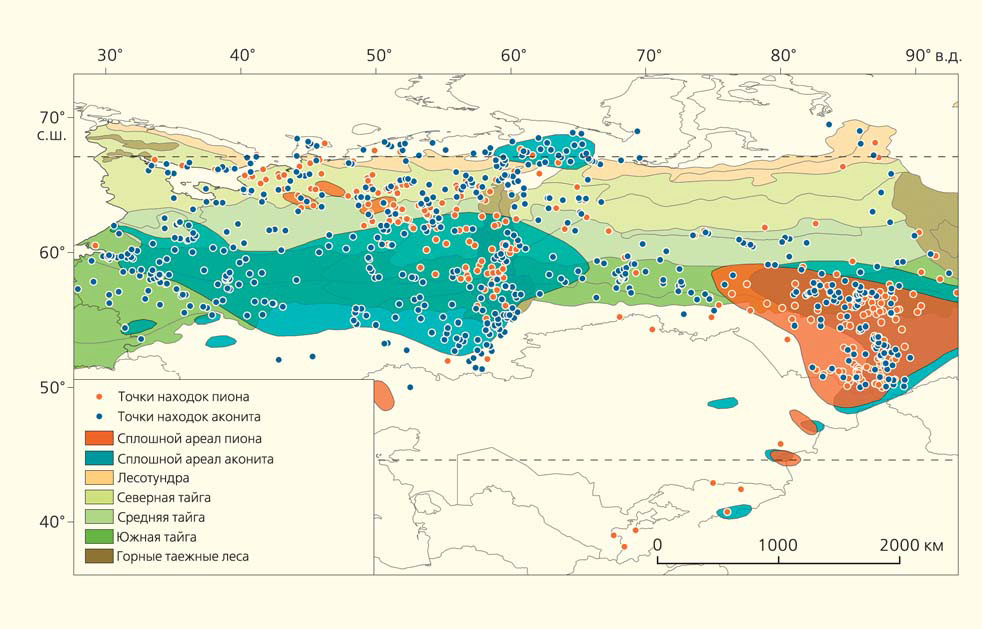
Ареалы пиона и аконита
Всего в высокотравных лесах насчитывается не менее 41 вида цветковых растений и четыре вида папоротников — такие данные приводятся в 850 имеющихся на сегодняшний день геоботанических описаниях. Сравнение видовых ареалов, взятых из базы «Ареал» (составлена Т. С. Проказиной по литературным и гербарным материалам), приводит к выводу, что большинство видов — 31 из 45 проанализированных — произрастает в лесах всей Северной Евразии. Это свидетельствует о широчайшем распространении высокотравных лесов в прошлом и служит основанием для их более широкого поиска на востоке России.
Второй ярус напочвенного покрова этих лесов также необычен. Здесь растут неморальные травы, по своему происхождению связанные с широколиственными лесами. Это ранневесенние (т. е. эфемероиды): хохлатка плотная (Corydalis bulbosa), гусиный лук (Gagea lutea), ветреница алтайская (Anemone altaica). Кроме них — летневегетирующие: сочевичник (Lathyrus vernus), бор (Milium effusum), вороний глаз (Paris quadrifolia) и звездчатка (Stellaria holostea).

Эфемероиды хохлатка плотная, гусиный лук желтый, ветреница алтайская на поляне в высокотравном лесу весной. Печоро-Илычский заповедник. Фото А. С. Ефименко
Неморальные виды, а из них особенно эфемероиды, вся надземная жизнь которых проходит до формирования лиственного полога и требует достаточно света, в распространении тесно связаны с широколиственными деревьями и осиной. В темнохвойной же тайге присутствие этих видов выглядит неожиданным. Однако реконструкция ареалов широколиственных пород в позднем голоцене приводит к выводу, что они распространялись значительно дальше на север, чем в настоящее время [13]. Вероятно, неморальные виды в темнохвойных лесах подтверждают это. Надо особо отметить, что эфемероиды в тайге сохранились только в высокотравных лесах с их богатыми и влагоемкими почвами, и преимущественно на полянах, где света вполне достаточно.
В третьем ярусе обитают мелкие травы: кислица, седмичник, майник и другие, которые растут и в зеленомошных лесах.
Четвертый ярус образуют стелющиеся по поверхности почвы гемибореальные мхи рода Plagiomnium, а также Barbilophozia lycopodioides, Brachythecium reflexum, Rhizomnium magnifolium, Rhodobryum roseum и др. В зеленомошных лесах эти виды редки. Характерные же для зеленомошных лесов мхи, такие как Pleurozium schreberi и Hylocomium splendens, на почве высокотравных не растут. Здесь их единственное местообитание, где они доминируют, — валежины, на которых в начальных стадиях зарастания эти мхи формируют сплошной ковер (в небольшом количестве также имеются виды бореального мелкотравья и черника).

Высокотравный темнохвойный лес в фазу цветения пионов. Печоро-Илычский заповедник. Фото А. А. Алейникова
Есть еще одна удивительная особенность высокотравных лесов. В основании стволов крупных елей и кедров растет брусника и встречаются кустистые лишайники родов Cladonia и Cladina — типичные обитатели сосняков. Невероятное множество эколого-ценотических групп (видов, предпочитающих сходное местообитание) в напочвенном покрове высокотравных лесов — результат большого разнообразия условий обитания (микроместообитаний) растений. Так, кроме валежин в высокотравных лесах обычны западины — ямы и бугры, формирующиеся в результате выворачивания кома земли падающим деревом. В ямах часто поселяются нитрофильные растения, которые хорошо растут только на почвах, богатых биодоступными соединениями азота. Среди этих обычных обитателей приречных ольховых лесов — селезеночник очереднолистный (Chrysosplenium alternifolium), звездчатка дубравная (S. nemorum), таволга вязолистная (Filipendula ulmaria). А на буграх растут лугово-опушечные виды — вика (Vicia sepium), мятлик (Poa pratensis) и др. По мере разложения валежин, зарастания бугров и западин, т. е. по мере изменения микроместообитаний, меняется и видовой состав растений: на смену специфическим видам приходят фоновые. Поскольку валежины, бугры и западины появляются постоянно, в высокотравных темнохвойных лесах одновременно существует огромное разнообразие экологически различных видов сосудистых растений и мхов [14].

Элементы ветровально-почвенного комплекса: бугор и западина в высокотравном лесу. Печоро-Илычский заповедник. Фото А. А. Алейникова

Бурозем грубогумусный в высокотравном лесу. Печоро-Илычский заповедник. Фото А. Д. Бовкунова
Самые обычные почвы высокотравных лесов — буроземы типичные и грубогумусные. Их диагностические признаки: хорошо структурированный гумусовый горизонт мощностью до 30–60 см (иногда более), относительно высокое содержание органического вещества (3,5–5,5%), нейтральный или слабокислый рН (от 5,0 до 6,2), хорошее обеспечение кальцием, калием и фосфором по сравнению с другими почвами таежных лесов.
Почвы высокотравных лесов высоковлажные, поэтому снижена возможность их возгорания. На водоразделах в большинстве почвенных профилей признаки пожаров не обнаружены [14]. Сравнение растительности и почвы двух типов северных лесов — зеленомошных и высокотравных — свидетельствует о влиянии антропогенного фактора на развитие и формирование современной тайги, меняет наше представление о ее природном облике.
Индикаторы состояния лесов
К настоящему времени в восточноевропейских заповедниках накоплен большой материал по динамике лесов. Подобраны ряды растительных сообществ, живущих в сходных экотопических (внешних) условиях, но отличающихся давностью и типами антропогенных нарушений. Эти исследования позволили выделить набор признаков-индикаторов сукцессионного (от лат. successio — ‘преемственность’, ‘наследование’) состояния лесных сообществ:
- соотношение раннесукцессионных (пионерных) и позднесукцессионных (климаксовых) видов, вычисленное по сумме площадей поперечных сечений деревьев;
- распределение позднесукцессионных видов деревьев по возрастному состоянию (по типам онтогенетических спектров);
- наличие ветровальных почвенных комплексов: бугров, валежин и западин;
- разнообразие эколого-ценотической структуры напочвенного покрова;
- видовое богатство (число видов, характерное для сообщества) и видовая насыщенность (число видов на единицу площади) сосудистых растений и мхов.
Процессы восстановления природного облика лесов (аутогенные сукцессии) характеризуются определенными закономерностями в изменении признаков-индикаторов. Анализ рядов лесных сообществ, выстроенных по давности антропогенных нарушений, позволил нам выявить пять следующих закономерностей.
- Сумма площадей поперечных сечений деревьев пионерных видов на начальных стадиях аутогенной сукцессии близка к 100%, а на завершающих стадиях их количество снижается, при этом доля климаксовых не превышает 15%.
- Популяции деревьев климаксовых видов на среднем этапе сукцессии представлены только семенами или молодыми особями, а на завершающем — это нормальные полночленные популяции, что говорит об устойчивом потоке поколений.
- Системы микроместообитаний — бугры, западины и валежины — на среднем этапе сукцессии в сообществе только формируются, а на завершающем присутствуют во всех стадиях преобразования примерно в равных соотношениях.
- В напочвенном покрове после смыкания полога темнохвойных деревьев в начале развития сообщества произрастают виды одной-двух эколого-ценотических групп, на завершающих стадиях — виды всех групп или их большинства. Последнее обстоятельство отражает максимально возможное природное разнообразие микроместообитаний.
- Видовое богатство и видовая насыщенность в начале сукцессии минимальны, а на поздних ее стадиях — максимальны для данной территории. Таким образом, благоприятно сочетаются множество различных местообитаний и наличие семян и спор всех видов растений, способных заселить эти места.
Именно высокотравным лесам присуща максимальная выраженность перечисленных признаков.
Онтогенетические спектры ели и пихты в этих сообществах нормальные, полночленные. Здесь есть полный набор бугров, западин и валежин, свидетельствующий о постоянном поддержании гетерогенности высокотравных лесов [15, 16]. Видовое богатство сосудистых растений в разных частях изученной территории, представленной на карте ареалов, достигает 150–200 видов, видовая насыщенность — 30–60 видов на 100 м2. По суммарному покрытию и фитомассе в напочвенном покрове доминирует высокотравье. В зеленомошных лесах все эти показатели значительно снижены. Видовое богатство сосудистых растений составляет 30–50 видов, видовая насыщенность — 5–10 видов. По суммарному покрытию и фитомассе в напочвенном покрове доминируют зеленые мхи [6, 14].
Итак, оценка по признакам-индикаторам состояния высокотравных темнохвойных лесов позволила заключить, что они находятся на завершающем этапе восстановления после нарушений, в то время как зеленомошные — на среднем. Судя по тому, что в почве большинства таежных лесов можно найти уголь, наиболее вероятная причина нарушений — мощные верховые пожары.
Давность последних пожаров (по предварительным радиоуглеродным датировкам) в высокотравных лесах Печоро-Илычского заповедника составляет 600–1000 и более лет, а в зеленомошных не превышает 200–250 лет.
Вполне вероятно, что часть исследованных высокотравных лесов не испытала мощных антропогенных воздействий и представляет собой рефугиумы (прибежища, укрытия) истинно природной тайги. Иными словами, возможно, это участки, избежавшие влияния человека. Чтобы четко различить обозначенные таким образом два варианта высокотравных лесов, необходимы дополнительные исследования их биоразнообразия и структурно-функциональной организации.
Доисторический облик тайги
Полученные результаты позволяют реконструировать природный облик тайги. Можно предположить, что именно высокотравные леса определяли его. Об этом свидетельствуют их распространение от северных пределов лесного пояса до южной тайги, экотопическое разнообразие и способность существовать в суровом климате, сохраняя смешанный (неморально-бореальный) характер флоры.
Наше предположение созвучно учению о происхождении современной таежной зоны из арктотретичных (т. е. распространённых в миоцене по всей полярной области) лесов в результате постепенного изменения климата. Эти представления были независимо сформулированы А. И. Лесковым [17] и В. Б. Сочавой [18] на основе биогеографических исследований, а также на базе палеоботанических — В. П. Гричуком [19] и П. И. Дорофеевым [20]. Неморально-бореальный характер травяно-кустарничкового яруса высокотравных темнохвойных лесов также можно рассматривать как аргумент в пользу происхождения современной тайги из арктотретичных лесов [12, 21]. Такой ярус может сформироваться только при наличии: опада лиственных деревьев (поскольку под их пологом чаще встречаются неморальные виды); западин, где обычно обитают нитрофильные растения и гемибореальные мхи; валежин, на которых концентрируются бореальные мелкие травы, кустарнички и зеленые мхи; межкроновых участков, с господством высокотравья и др. Поэтому произрастание неморальных видов трав в северной и средней тайге, где есть только мелколиственные деревья, косвенно может свидетельствовать о былом присутствии в них и широколиственных.
Высокотравные леса ярко выделяются среди всех северных таежных лесов своим видовым богатством. Мы уверены, что это наиболее значимый объект для оценки биологического разнообразия тайги и возможностей реализации ее экосистемных функций.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (проект 13-04-01491а), программы Президиума РАН «Живые системы» и гранта Президента РФ (МК-2102.2012.4).
Литература
1. Curtis I. A. Valuing ecosystem goods and services: a new approach using asurrogate market and the combination of a multiple criteria analysis and a Delphi panel to assign weights to the attributes // Ecologic. Economics. 2004. V. 50. № 3–4. P. 163–194.
2. Горшков В. Г., Макарьева А. М., Лосев К. С. Планетарный парниковый эффект и биотическая устойчивость климата Земли // Изв. РАЕН. Секция наук о Земле. 2001. Вып. 7. С. 62–68.
3. Мониторинг биологического разнообразия лесов России: методология и методы / Ред. А. С. Исаев. М., 2008.
4. Толмачев А. И. К истории возникновения и развития темнохвойной тайги. М.; Л., 1954.
5. Смирнова О. В., Коротков В. Н. Старовозрастные леса Пяозерского лесхоза северо-западной Карелии // Ботан. журн. 2001. Т. 86. № 1. С. 98–109.
6. Смирнова О. В., Бобровский М. В., Ханина Л. Г., Смирнов В. Э. Сукцессионный статус старовозрастных темнохвойных лесов Европейской России // Успехи совр. биол. 2006. № 1. С. 26–48.
7. Бобровский М. В. Лесные почвы Европейской России: биотические и антропогенные факторы формирования. М., 2010.
8. Восточноевропейские леса: история в голоцене и современность / Ред. О. В. Смирнова. Кн. 1–2. М., 2004.
9. Сукцессионные процессы в заповедниках России и проблемы сохранения биологического разнообразия / Ред. О. В. Смирнова и Е. С. Шапошников. СПб., 1999.
10. Оценка и сохранение биоразнообразия лесного покрова в заповедниках Европейской России / Ред. Л. Б. Заугольнова. М., 2000.
11. Заугольнова Л. Б., Мартыненко В. Б. Определитель типов леса Европейской России. М., 2013.
12. Заугольнова Л. Б., Смирнова О. В., Браславская Т. Ю. и др. Высокотравные таежные леса восточной части Европейской России // Растительность России. 2009. № 15. С. 3–26.
13. Смирнова О. В., Бакун Е. Ю., Турубанова С. А. Представление о потенциальном и восстановленном растительном покрове лесного пояса Восточной Европы // Лесоведение. 2006. № 1. С. 22–33.
14. Смирнова О. В., Алейников А. А., Семиколенных А. А. и др. Пространственная неоднородность почвенно-растительного покрова темнохвойных лесов в Печоро-Илычском заповеднике // Лесоведение. 2011. № 6. С. 67–78.
15. Алейников А. А., Бовкунов А. Д. Микромозаичная организация крупнопапоротниковых и высокотравных пихто-ельников Печоро-Илычского заповедника // Изв. ПГПУ им. В. Г. Белинского. 2011. № 25. С. 21–35.
16. Алейников А. А., Лазников А. А. Пространственная организация крупнопапоротниковых лесов в верховьях реки Печоры (Печоро-Илычский заповедник) // Изв. Самарского научного центра РАН. 2012. Т. 4. № 1 (5). С. 1180–1183.
17. Лесков А. И. Принципы естественной системы растительных ассоциаций // Ботан. журн. 1943. Т. 28. № 2. С. 37–52.
18. Сочава В. Б. Вопросы флорогенеза и филоценогенеза маньчжурского смешанного леса // Материалы по истории флоры и растительности СССР. М.; Л., 1946. Вып. 1. С. 283.
19. Гричук В. П. История флоры и растительности Русской равнины в плейстоцене. М., 1989.
20. Дорофеев П. И. Новые данные о плейстоценовых флорах Белоруссии и Смоленской области // Материалы по истории флоры и растительности СССР. Вып. 4. М.; Л., 1963. С. 5–180.
21. Смирнова О. В., Калякин В. Н., Турубанова С. А., Бакун Е. Ю. Генезис восточноевропейской тайги в голоцене // Закономерности вековой динамики биогеоценозов. XXI чтения памяти академика В. Н. Сукачева. М., 2006. С. 18–65.
-
Высокотравные леса, имеющие в составе неморальные элементы, поддерживаются почвенным избыточным увлажнением. Если фактор переувлажнения убрать, эти леса быстро превратятся в ходе сукцессии в мелкотравно-кустарничково-зеленомошную тайгу. Все наоборот.
Как определить ранне- и позднесукцессионные виды без схемы сукцессии?










Общий вид зеленомошного ельника. Национальный парк «Паанаярви». Северная Карелия. Здесь и далее фото М. В. Бобровского