Взаимодействие растений с насекомыми: палеозойская история
Сергей Наугольных
«Природа» №3, 2017
 Об автореСергей Владимирович Наугольных — доктор геолого-минералогических наук, главный научный сотрудник Геологического института РАН, профессор РАН; контрактная аффилиация — ведущий научный сотрудник научно-исследовательской лаборатории стратиграфии Казанского федерального университета. Область научных интересов — палеоэкология, систематика и эволюция высших растений, изучение палеопочв. |
Над лесом сигиллярий, над лесом фавулярий
Летела голденбергия вперед.
А где-то там на ветке, а где-то там на вайе
Висел вполне созревший птеридосперма плод.
Подлетает она, подлетает, тихо крыльями шурша.
И съедает его, и съедает, сок вкушая не спеша.
Наземные растения, начиная с глубочайшего геологического прошлого, вовлечены в сложные взаимоотношения с самыми разными компонентами экосистем, в первую очередь с почвой, с соседями по растительному сообществу — фитоценозу, с грунтовыми водами и с атмосферными осадками. И практически с самого момента своего появления высшие растения находились и продолжают находиться в тесной взаимосвязи с различными членистоногими, прежде всего насекомыми.
Взаимодействие это было многовалентным (то негативным, то позитивным), и в конце концов оно привело к образованию симбиотической пары «опыляемый — опылитель», без появления которой, пожалуй, не было бы того чарующего разнообразия покрытосеменных растений, которое окружает нас сегодня.
Давайте попробуем взглянуть на ранние этапы коэволюции растений и насекомых, для того чтобы попытаться ответить на один из традиционных вопросов в теории эволюции: чего в природе больше — ожесточенной борьбы за существование или, напротив, симбиотического сотрудничества?
Питание
Вне зависимости от того, кто мы — убежденные вегетарианцы или, наоборот, любители разных гастрономических изысков, на нашем столе всегда находится большое количество продуктов растительного происхождения. Именно растения дают нам углеводы, обеспечивающие энергетику организма, и многие из белков, позволяющих строить и обновлять мышечную массу. Нужны и растительные жиры (вспомним хотя бы подсолнечное масло), способствующие образованию запасов питательных веществ «на черный день».
Именно с трофической точки зрения растения рассматривались насекомыми с первых же дней их совместной эволюции в наземных экосистемах. Известны многочисленные и разнообразные краевые погрызы листовых пластинок, оставленные насекомыми и многоножками-диплоподами на листьях палеозойских птеридоспермов. В середине пермского периода насекомые (а именно жуки пермокупедиды) научились переваривать и усваивать целлюлозу древесины древних голосеменных — кониферофитов [1].
Однако у растений нашлись и более лакомые для насекомых части — спорангии, а позднее и семязачатки, насыщенные углеводами и белками.

В каменноугольных и пермских отложениях Ангариды (древнего материка, располагавшегося на месте северо-восточной части современной Евразии) встречаются семена с маленькими круглыми отверстиями — следами проникновения длинных челюстей-стилетов внутрь семязачатка, через которые насекомое высасывало его вкусное содержимое — так же, как гурман пьет через трубочку коктейль. Пример таких насекомых — голденбергия (Goldenbergia), описанная из нижнепермских отложений Сибири. Голосеменных растений в то время в этом регионе уже было множество, и они еще не научились защищаться от ловких любителей отведать палеозойского аналога кокосового молока.
В пермских отложениях Приуралья время от времени попадаются семена, описанные как Cardiocarpus cordatus. На некоторых из них можно обнаружить аккуратные дырочки, расположенные почему-то всегда у самого основания семени. Не было понятно, каким именно растениям принадлежали эти семена. Кто сделал дырочки, тоже оставалось неизвестным. Выдающийся российский палеонтолог А. Г. Шаров высказал предположение, что похожие перфорации были оставлены на семенах из верхнепалеозойских отложений Сибири палеодиктиоптерами [2]. Представители палеодиктиоптер — насекомые рода парадунбария (Paradunbaria) — встречаются в нижнепермских отложениях Приуралья, где были найдены семена кардиокарпуса кордатуса с перфорациями. Судя по всему, именно парадунбария и высасывала содержание семязачатков C. cordatus. Но почему дырочки от проколов располагаются в основании семян? Ответ подсказали исследования Поташки — интереснейшего местонахождения ископаемых животных и растений, расположенного в Артинском р-не Свердловской обл.

Семя Cardiocarpus cordatus с проколом в основании (показан стрелкой) из отложений артинского яруса нижней перми Приуралья (местонахождение Сухановка-1)
Строго говоря, Поташка — это не одно местонахождение, а целый комплекс из четырех местонахождений, получивших названия Поташка-1, -2, -3 и -4, а также находящихся по соседству местонахождений Сухановка-1 и -2. В них обнажаются зеленоватые полимиктовые песчаники артинского яруса нижнего отдела пермской системы. Эти же песчаники выходят на дневную поверхность и к северу — у г. Арти на горе Кашкабаш, где находится исторический стратотип артинского яруса. Их, собственно, и изучал академик А. П. Карпинский, установивший артинский ярус, который с той поры частенько называют ярусом Карпинского. Сами песчаники характерного буровато-зеленого оттенка нередко называют жерновыми, поскольку из них выпиливали большие круглые жернова для сельских мельниц.
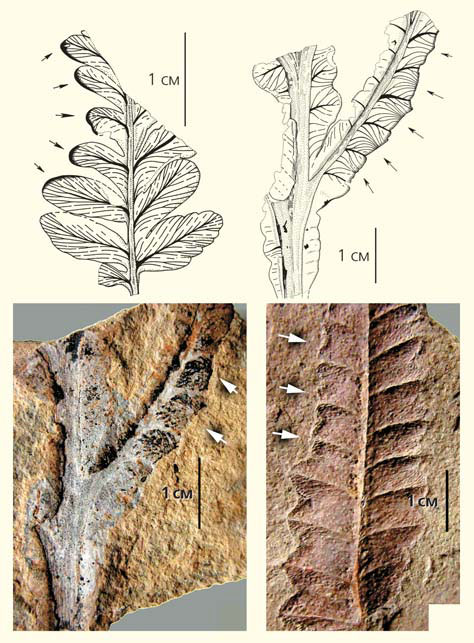
Листья пельтаспермового птеридосперма Permocallipteris retensoria с краевыми погрызами (отмечены стрелками) из отложений кунгурского яруса нижней перми Приуралья (местонахождение Чекарда-1)
В целом жерновые песчаники довольно бедны органическими остатками. В них время от времени попадаются раковины головоногих моллюсков аммоноидей, которым Карпинский посвятил одну из своих блестящих палеонтологических работ. Помимо аммоноидей в отдельных прослоях и в особенности в конкрециях можно найти растительные остатки хорошей сохранности. Из растений здесь встречаются древние хвощевидные рода Paracalamites, отдельные перья папоротников, листья пельтаспермовых птеридоспермов, гинкгофитов и войновскиевых, побеги хвойных и разнообразные семена. Однако в количественном отношении во всех четырех Поташках совершенно отчетливо доминируют семена C. cordatus и декортицированные побеги вальхиевых хвойных. Облиственные побеги, непосредственно относящиеся к роду Walchia, тоже время от времени встречаются в этих местонахождениях, но реже.
Для меня после изучения Поташки стало совершенно очевидно, что C. cordatus — это семена вальхиевых хвойных. Но эта группа растений в отложениях нижней перми Приуралья представлена несколькими видами. К какому именно из них принадлежали эти семена, еще предстояло установить.
Проблема в том, что семена голосеменных в целом и хвойных в частности довольно редко сохраняются прикрепленными к семеносным органам, поэтому задачу предстояло решить формально-логическим путем. Исключив из рассмотрения всех кандидатов, заведомо не подходивших на роль растений, которые продуцировали семена C. cordatus, я остановил свой выбор на двух видах — Walchia appressa и W. bardaeana. Оба этих вида вальхиевых хвойных были описаны знаменитым российским палеоботаником М. Д. Залесским еще в 1937 г. [3]. Ему было неизвестно, как были устроены семеносные органы этих хвойных, но, поскольку их облиственные побеги различались довольно отчетливо, никаких сомнений в том, что это два разных самостоятельных вида, ни тогда, ни сейчас не возникало и не возникает.
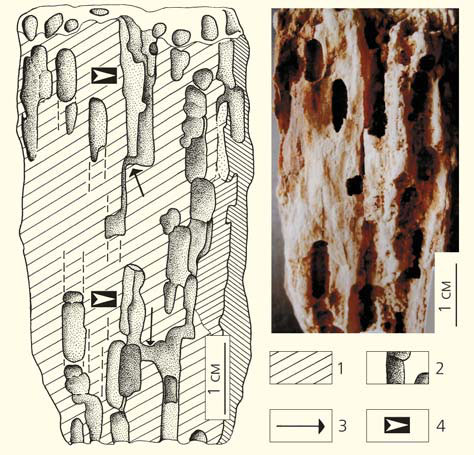
Древесина кониферофита Dadoxylon sp. с ходами жуков из семейства пермокупедид из отложений казанского яруса средней перми Прикамья (местонахождение Тихие Горы): 1 — продольное сечение фрагмента древесины; 2 — полости в древесине, 3 — тоннели, 4 — шахты
Женские стробилы вида W. appressa значительно позднее были найдены и описаны С. В. Мейеном, другим ярким российским палеоботаником и эволюционистом. Найденные стробилы ученый описал под собственными родовым и видовым названиями Kungurodendron sharovii [4]. Но после обнаружения стробилов этого типа в прикреплении к облиственным ветвям W. appressa, которые были обнаружены в еще одном знаменитом местонахождении ископаемых животных и растений пермского возраста — Чекарде [5], стало совершенно очевидно, что это один и тот же вид исходного растения.
Семенная чешуя W. appressa имеет отчетливо радиально-симметричное строение. Семенная чешуя несет не менее восьми-девяти мелких изогнутых семяножек. Семена C. cordatus на них размещаться не могли — просто не хватило бы места в пазухе брактеи (стерильного придатка чешуевидной формы, прикрывающего семенную чешую) стробила. Таким образом, W. appressa из дальнейшего рассмотрения была исключена.
Надежно установить, какими были женские стробилы другого вида вальхиевых хвойных — W. bardaeana, долгое время не удавалось, пока во время полевых работ моим коллегам из Кунгурского историко-архитектурного и художественного музея-заповедника не улыбнулась удача. В местонахождении, расположенном в Пермском крае, недалеко от с. Матвеева, на правом берегу р. Барды, им удалось найти побег W. bardaeana с женским репродуктивным органом хорошей сохранности. К моему удивлению, этот орган оказался совсем не стробилом, а фертильной зоной, т. е. участком побега с располагающимися на нем брактеями и семенными чешуями. Выше этого фертильного участка находился обычный побег с нормальными листьями. Он к тому же ветвился.
Внимательно рассмотрев находку, я внезапно осознал, что похожая фертильная зона уже есть в моей коллекции. Стерильные листья на ней не сохранились, из-за чего нельзя было установить, что этот орган принадлежал W. bardaeana. Я даже собирался описать имевшийся у меня экземпляр как новый вид.

Ископаемые остатки вальхиевого хвойного Walchia bardaeana из отложений артинского (2, 3) и кунгурского (1, 4, 5) ярусов нижней перми Приуралья: 1 — средняя часть женской фертильной зоны (местонахождение Красная Глинка), 2, 3 — семя Cardiocarpus cordatus, стрелкой отмечен семенной рубчик — место, посредством которого семязачаток прикреплялся к семяножке (местонахождение Актюбинск); 4, 5 — облиственные побеги W. bardaeana (местонахождение Таежное-1)
После детального изучения фертильных зон этого вида удалось установить, что в их брактейно-пазушных комплексах имелась единственная уплощенная семяножка, окруженная многочисленными игловидными или чешуевидными стерильными придатками. В верхней части семяножки на ее стороне, обращенной к оси фертильной зоны, находился семенной рубец, к которому при жизни растения прикреплялось единственное семя. Расширяющаяся кверху семяножка своей формой напоминала очертания семян C. cordatus. Семя, очевидно, должно было прикрепляться к семяножке только одной стороной. Иными словами, семенной рубчик на семенах C. cordatus должен был быть смещен на одну из сторон семени. Внимательно рассматривая наиболее хорошо сохранившиеся семена этого вида, имеющиеся в моей коллекции, я обнаружил, что так оно и есть! Раньше я этого не замечал.

Парадунбария на фертильной зоне Walchia bardaeana. Ранняя пермь. Приуралье
Стало понятно, что семена C. cordatus сидели на семяножках обращенно, инвертированно — своей микропилярной частью (т. е. верхушкой семени) вниз к оси фертильной зоны. Наружу выглядывало только основание семени. Именно его и протыкали своим хоботком-стилетом парадунбарии. Вот почему дырочки от их проколов располагаются в основании семян! На основе всех этих наблюдений и умозаключений я подготовил графическую реконструкцию фертильной зоны W. bardaeana с сидящей на ней парадунбарией.
Микроперфорации, сходные с дырочками на семенах C. cordatus, но более мелкие, были обнаружены на стенках спорангиев мужских фруктификаций рода пермотека (Permotheca). Органы размножения этого типа были характерны для пельтаспермовых (о них подробнее будет рассказано ниже) и ранних гинкгофитов. Наличие таких перфораций на стенках спорангиев пермотек указывает на то, что и они вызывали трофический интерес у насекомых (скорее всего, все тех же палеодиктиоптер).

Мужские фруктификации Permotheca spp. из отложений кунгурского яруса нижней перми Приуралья (местонахождение Чекарда-1): 1 — макроморфологическое строение синангия; 2 — реконструкция средней части кистевидного собрания синангиев; 3, 4 — прокол стенки спорангия
Защита
Раз есть угроза для семян, иными словами — для потомства, значит, надо искать выход из складывающейся драматической ситуации. Как говорится, «эволюционируй или вымирай». Развитие мощных покровных чешуй у хвойных (вспомним женские шишки современных елей и сосен) было вполне адекватным и эффективным ответом на эту угрозу.
Другая группа палеозойских голосеменных растений — птеридоспермы — пошла по иному пути. Их семена изначально сидели на листовидных органах, модифицированных в различной степени. Семена, таким образом, были открыты для всех желающих подзакусить из мира насекомых (например, для всё той же голденбергии). Подлетай, питайся!
Но уже вскоре птеридоспермы «научились» защищаться. Края фертильных листьев (мегаспорофиллов) с сидящими на них семенами стали подворачиваться, прикрывая семязачатки, причем этот процесс, по всей видимости, происходил независимо в разных группах птеридоспермов.
Из растений, произраставших в пермском периоде в Ангариде, и в особенности на тех территориях, которые спустя 270 млн лет стали Приуральем, в этом отношении весьма примечательны пельтаспермовые птеридоспермы порядка Peltaspermales. Таксономические границы этого порядка трактуются разными исследователями существенно по-разному. По моему мнению, объем этого порядка следует ограничить только тремя семействами: Peltaspermaceae, Angaropeltaceae и Vetlugospermaceae [6].
Женские репродуктивные органы растений, относящихся к семейству Peltaspermaceae, выглядели как кистевидные собрания семеносных дисков. Каждый семеносный диск (их еще называют пельтоидами) состоял из округлого уплощенного мегаспорофилла, к центру которого с нижней (адаксиальной) стороны прикреплялась ножка. Диск обычно был разделен на радиальные сектора, количество которых равнялось числу семян. Края диска могли быть ровными, фестончатыми или лопастными. Концентрически вокруг ножки к адаксиальной (т. е. обращенной к оси, к побегу) поверхности диска прикреплялись семена. Мегаспорофиллы (пельтоиды) посредством ножки соединялись с осью предыдущего порядка так, что все собрание мегаспорофиллов, по существу, становилось стробилом, шишкой. До созревания семян края соседних щитков (дисков) могли смыкаться, а после созревания края расходились, семена отпадали от мегаспорофиллов и падали вниз под действием силы тяжести, поскольку были лишены крылатки.
Однако у представителей эволюционно более продвинутого семейства ангаропельтовых (Angaropeltaceae) возникло важное адаптивное новшество. Края мегаспорангиатных дисков этих растений загнулись вниз к ножке пельтоида до такой степени, что при этом образовалась практически полностью закрытая капсула с семенами внутри [7]. Между краем диска и ножкой пельтоида осталась лишь узкая щель. В нее высовывались микропилярные части семян, а вот зачем — поговорим отдельно.
То, что этот процесс преобразования семеносного диска в капсулу был вызван необходимостью защитить семена от неблагоприятного воздействия негативных факторов среды, — в общем-то, очевидно. Но до какой степени это могло быть важно для растения, удалось понять только после изучения одного из родов ангаропельтовых — сылвокарпуса (Sylvocarpus).
Семеносные капсулы этого растения были устроены относительно просто. Ножка, как и у всех остальных настоящих пельтаспермовых (порядок Peltaspermales), прикреплялась к центральной части адаксиальной поверхности капсулы. Края капсулы изгибались книзу, практически примыкая к ножке. Очень похоже на то, что до созревания семян даже небольшая щель между ножкой капсулы и краем мегаспорофилла тоже была прикрыта защитной пленкой. В пространстве под щитком капсулы находилось шесть семян. Первоначально предполагалось, что у голотипа их было только пять, однако дополнительное препарирование образца и находки новых экземпляров позволили установить, что и у типового экземпляра, и практически у всех других представителей этого вида было по шесть семян в капсуле.
Верхняя поверхность капсулы была покрыта необычно толстой кутикулой. Я изучил ее под электронным сканирующим микроскопом и был вознагражден за усердие. Оказалось, что на кутикуле сохранились явные следы округлых перфораций, оставленные каким-то насекомым, которое пыталось добраться до семян, заключенных внутри капсулы. Однако сделать это насекомому не удалось. Кутикула, покрывавшая капсулу, была настолько прочной и толстой, что ни один из проколов не оказался сквозным. Насекомому просто не удавалось преодолеть надежную защиту.
Я назвал этот новый вид сылвокарпус арматус (Sylvocarpus armatus), т. е. «сылвенский плод защищенный». Думаю, это один из самых первых примеров в эволюционной истории растений, когда в бесконечной «гонке вооружений» растениям удалось выработать надежный механизм защиты от непосредственного нападения.

Семеносные капсулы Sylvocarpus armatus из отложений кунгурского яруса нижней перми Приуралья (1, 3–5 — местонахождение Чекарда-1, 2 — Таежное-1): 1, 2 — макроморфология; 3–5 — перфорации в кутикуле
Опыление
Настало время вспомнить об узкой щели между краем и ножкой семеносной капсулы ангаропельтовых. Зачем же она была нужна?
Для оплодотворения семязачатка у голосеменных необходимо, чтобы на верхушку семени, где располагается микропилярный канал, попала пыльца, которая затем самостоятельно или прилипнув к капле вязкой жидкости, выделяемой семязачатком, проникает в пыльцевую камеру. Далее следует собственно процесс оплодотворения семязачатка и развития эмбриона нового растения.
Пока семязачатки просто висят на листе, как у относительно примитивных каменноугольных птеридоспермов, никаких особых проблем с их оплодотворением нет. Ветер носит по воздуху массу пыльцы, которая без особых сложностей попадает в микропиле семязачатков. Но вот если семязачатки заключены внутрь капсулы, пыльце проникнуть в микропиле не так-то просто.
На поверхности семеносных органов у представителей всех трех семейств пельтаспермовых (и Peltaspermaceae, и Angaropeltaceae, и Vetlugospermaceae) имелись миниатюрные секреторные органы — гланды, которые, возможно, выделяли феромоны, привлекавшие насекомых-опылителей. Считать это твердо доказанным пока нельзя, но иначе зачем тогда эти секреторные органы?
Мы с А. А. Оскольским, моим коллегой из Ботанического института РАН, рассудили, что гипотеза о насекомоопылении, или энтомофилии, пельтаспермовых вполне имеет право на существование, и посвятили этой проблеме специальную статью, опубликованную на страницах международного ботанического журнала Wulfenia [8].
Мимикрия
Конечно же, не надо забывать, что и между самими насекомыми в лесах позднего палеозоя велась жестокая и напряженная борьба за существование. Правила этой борьбы были просты: если атакуешь, то будь быстрее и сильнее жертвы, если защищаешься — создавай плотные надежные покровы (панцирь) или прячься так, чтобы не нашли.
Похоже, именно в пышных лесах каменноугольного периода у насекомых впервые появилась новая стратегия выживания: маскироваться под листья или кору процветавших в карбоне споровых и голосеменных растений. Эта мода была подхвачена и продолжена насекомыми пермского периода.
Давайте посмотрим на жилки крыльев и надкрыльев пермского таракана. Тараканы того времени были под стать современным мадагаскарским, а то и крупнее. Жилки на тараканьем крылышке любому палеоботанику покажутся знакомыми, и неудивительно, ведь они почти точь-в-точь повторяют жилкование перышек ваий пельтаспермовых из формально-морфологической группы каллиптерид. Чтобы укрыться от зорких глаз хищника, таракану было достаточно замереть на листе пермокаллиптериса (Permocallipteris), и происходило чудо: взор хищника равнодушно, ничего не замечая, скользил по растению, на котором играли солнечные блики и тени ветвей.

Листья каллиптерид и позднепалеозойские тараканы: 1 — крыло таракана из пермских отложений Печорского угольного бассейна, 2 — перистый лист пельтаспермового Permocallipteris retensoria, 3 — позднепалеозойский таракан Phylloblatta на вайе птеридосперма (реконструкция таракана по [12]); 4, 6 — таракан Kunguroblattina microdictya из нижнепермских отложений Приуралья (по [13]), 5 — перышко (сегмент последнего порядка) пельтаспермового Permocallipteris adzvensis из среднепермских отложений (казанский ярус) Печорского угольного бассейна
А хищников, готовых полакомиться вкусным сочным тараканом, в лесах пермского периода было полным-полно. Это и амфибии, мелкие и крупные, чувствовавшие себя в конце палеозойской эры на суше не менее уверенно, чем в воде; и рептилии, стремительно перебегавшие из тени в тень в поисках добычи; и даже свои же собратья — насекомые, например гигантские хищные стрекозы меганевриды.
На мелководье пермских озер и лагун покачивались над водой побеги клинолистников сфенофиллов, а по ним то тут, то там ползали скорпионницы петромантисы. Им можно было чувствовать себя вполне спокойно: ведь когда они расправляли крылья в стороны, получалась точная копия розетки листьев сфенофилла, и никакой хищник их не замечал.

Мимикрия пермских скорпионниц: 1 — розетка листьев Sphenophyllum comiense из пермских отложений Печорского бассейна, 2 — Petromantis sylvaensis на побеге Sphenophyllum biarmicum (морфология петромантиса дана по [14]), 3 — розетка листьев S. biarmicum из пермских отложений Приуралья, 4 — облиственный побег S. comiense из пермских отложений Печорского бассейна
В палеонтологии известны счастливые случаи находок насекомых с сохранившейся окраской крыльев. Удивительно, но таких случаев не так уж и мало. Вот, например, великолепно сохранившийся отпечаток дунбарии (Dunbaria), родственницы нашей парадунбарии. Ее нашли в нижнепермских отложениях Канзаса [9]. На крыльях этого интересного насекомого прекрасно видны широкие темные поперечные полосы, соответствующие прижизненной пигментации. Зачем эти полосы были нужны?
Конечно же, это не мимикрия в полном смысле слова, но, тем не менее, тоже пример ловкой и эффективной маскировки. Это так называемая расчленяющая окраска. Вспомним камуфляж маскхалатов разведчиков, пятна на шкуре леопарда или полосы на шкуре зебры или тигра. Все они служат одной и той же цели: помочь животному спрятаться, стать незаметным. Одним животным это требуется для того, чтобы незаметно подкрасться и напасть, другим — чтобы не быть обнаруженным хищником.
Из желающих остаться незаметными можно вспомнить еще одну группу пермских насекомых гриллоблаттид. В лесах пермского периода жили представители рода тильярдембия (Tillyardembia). Этим насекомым яркая окраска была совершенно ни к чему. Тильярдембии, по мнению палеоэнтомологов, обитали в лесной подстилке, ползая среди мха и опавших листьев.
Яйцекладки
Проблема индивидуального выживания в животном и растительном мире тесно смыкается с не менее важной проблемой размножения. Что толку, если ты выжил здесь и сейчас, но не оставил потомства?
И здесь насекомые и растения вновь оказываются рядом. Яйца, отложенные кем-то из собратьев-насекомых, любой хищник воспримет как щедрый подарок судьбы. Еще бы — бесплатный лакомый белок безо всякой охраны подали вам буквально на блюдечке! Поэтому задача — сохранить потомство и надежно спрятать яйцекладку куда-нибудь подальше, но так, чтобы личинки не задохнулись и не погибли, — стала для многих первоочередной.
Но куда же спрятать яйцекладку? Одно из наиболее подходящих укромных мест — желобок в рахисе листа (т. е. в осевой его части, продолжающей черешок) крупного папоротника или птеридосперма, желательно поближе к основанию, подальше от пытливых глаз голодного хищника. Или, как вариант, прикопать яйца в почву, слегка присыпав их растительной трухой и перегноем. И надежно, и просто, и тепло: гниющие растения согревают яйца, помогая им развиваться. Но лучше всего, конечно же, отложить яйца на побег какого-нибудь растения, до которого никто не доберется...
Несколько лет назад мы сплавлялись по очень живописной уральской реке Сылве. По ее берегам, покрытым дремучей тайгой с вековыми елями, то тут, то там расположены обнажения терригенных пород пермского возраста, содержащих самые разнообразные и нередко хорошо сохранившиеся ископаемые остатки животных и растений.
Сделав одну из остановок у деревни Урма, мы прошлись вдоль берега Сылвы, приглядываясь к выходам песчаников артинского яруса. Вскоре я сделал замечательную находку: мощный, около 5 см в диаметре, и довольно длинный фрагмент побега, да даже и не побега, а ствола древнего представителя хвощевидных — паракаламита (Paracalamites). Внимательно осмотрев образец, я пришел уже в полнейший восторг: на поверхности ствола паракаламита были прекрасно видны огромные яйца, отложенные каким-то великаном из мира насекомых пермского периода. Долго гадать не пришлось: после работы с литературой и сравнения яйцекладок из Урмы с подобными находками из отложений другого возраста и из других регионов [10] удалось установить, что яйцекладка была оставлена гигантской стрекозой-меганевридой, скорее всего относящейся к роду арктотипус (Arctotypus). Отдельные крылья и даже целые арктотипусы время от времени встречаются здесь же — в пермских отложениях по берегам Сылвы и других уральских рек.

Стрекозы и их яйцекладки: 1 — крыло крупной стрекозы-меганевриды Arctotypus sp. из пермских отложений Приуралья (местонахождение Тюлькино); 2 — реконструкция стрекозы-меганевриды Arctotypus, откладывающей яйца на побеге паракаламита (по находке у д. Урма, р. Сылва, Пермский край); 3, 4 — яйцекладка стрекозы в желобке на рахисе птеридосперма из отложений кунгурского яруса нижней перми Приуралья (местонахождение Чекарда-1); 5 — яйцекладка стрекозы-меганевриды (отмечена стрелками) на стволе паракаламита из отложений артинского яруса нижней перми Приуралья (местонахождение Урма-2)
Очевидно, яйца были отложены на крупном паракаламите, ствол которого торчал из воды где-то на мелководье пермской лагуны или озера. Добраться до такой яйцекладки какому-нибудь наземному хищнику было просто невозможно.
***
Эпизоды из совместной истории палеозойских растений и насекомых, на мой взгляд, вполне красноречиво свидетельствуют в пользу прямой выгоды симбиотических взаимоотношений между различными компонентами экосистем даже в тех заведомо простых случаях, когда эти компоненты связаны только трофическими интересами. В том же случае, когда возникает такая форма симбиоза, как мутуализм, т. е. взаимовыгодное сотрудничество перед лицом бесконечных внешних угроз, возможности для выживания у обоих участников подобного союза резко возрастают. Хороший пример нам, людям, для решения насущных проблем в нашем непростом современном мире.
Работа выполнена в рамках исследовательских проектов Казанского (Приволжского) федерального университета в целях повышения его конкурентоспособности среди ведущих мировых научно-образовательных центров.
Литература
1. Наугольных С. В., Пономаренко А. Г. Предполагаемые следы питания жуков в древесине кониферофита из казанского яруса Прикамья // Палеонтологический журнал. 2010. № 4. С. 105–110.
2. Шаров А. Г. Морфологические особенности и образ жизни палеодиктиоптер // Докл. на двадцать четвертом ежегодном чтении памяти Н. А. Холодковского. Вопросы палеонтологии насекомых. Л., 1973. С. 49–63.
3. Zalessky M. D. Sur la distinction de l’étage Bardien dans le Permien de l’Oural et sur sa flore fossile // Problems of Paleontology. 1937. V. 2–3. P. 37–101.
4. Meyen S. V. Permian conifers of Western Angaraland // Rev. Palaeobot. Palynol. 1997. V. 96. P. 351–447.
5. Наугольных С. В. Флора кунгурского яруса Среднего Приуралья. М., 1998 (Тр. Геологического ин-та РАН. Вып. 509).
6. Naugolnykh S. V. Vetlugospermum and Vetlugospermaceae: a new genus and family of peltasperms from the Lower Triassic of Moscow syneclise (Russia) // Geobios. 2012. V. 45. P. 451–462.
7. Мейен С. В. Систематика пельтаспермовых птеридоспермов и их место в филогении голосеменных // Бюлл. МОИП. Отд. биол. 1983. Т. 88. № 1. C. 3–14.
8. Naugolnykh S. V., Oskolski A. A. An advanced peltasperm Permoxylocarpus trojanus Naug. from the Lower Permian of the Urals (Russia): an ancient case of entomophily in gymnosperms? // Wulfenia. 2010. V. 17. P. 29–43.
9. Dunbar K. O. Historical geology. NY; L., 1960.
10. Vasilenko D. V., Rasnitsyn A. P. Fossil ovipositions of dragonflies: review and interpretation // Paleontological Journal. 2007. V. 41. № 11. P. 1156–1161.
11. Историческое развитие класса насекомых / Ред. Б. Б. Родендорф, А. П. Расницын. М., 1980.
12. Guthörl P. Das Leben in den Süsswasserbecken und anderen Strand während der Rotliegendzeit im Saar-Nahe-Pfalz-Gebiet // Saarbrücker Bergmannskalender. 1948. S. 45–64.
13. Вишнякова В. Н. Новые виды рода Kunguroblattina из нижней перми Приуралья // Палеонтологический журнал. 1965. № 4. С. 50–59.
14. Novokshonov V. G. New and little known Permian scorpion flies (Insecta, Mecopteria = Panorpida) of the family Agetopanorpidae from Perm and Arkhangel’sk oblasts // Paleontological Journal. 1993. V. 27. № 1A. P. 70–89.












Goldenbergia sp., пролетающая над лесом раннепермской эпохи в Ангариде (морфология голденбергии дана по [11], с изм.). Здесь и далее рисунки автора