Есть ли почки у иглокожих?
Владимир Малахов, Ольга Ежова
«Природа» №7, 2016
 Владимир Васильевич Малахов — член-корреспондент РАН, заведующий кафедрой зоологии беспозвоночных биологического факультета МГУ им. М. В. Ломоносова, руководитель лаборатории биологии морских беспозвоночных Дальневосточного федерального университета (Владивосток). Область научных интересов — сравнительная анатомия, эмбриология и филогения беспозвоночных. |
 Ольга Владимировна Ежова — кандидат биологических наук, доцент той же кафедры, сотрудник той же лаборатории. Специалист в области морфологии и тонкой организации кишечнодышащих, морских звезд, офиур и голотурий; занимается изучением эволюции амбулакралий, морфологии, микроскопической анатомии и ультраструктуры полухордовых, иглокожих и хордовых. |
Иглокожие (Echinodermata) — морские звезды (Asteroidea), морские ежи (Echinoidea), офиуры, или змеехвостки (Ophiuroidea), голотурии (Holothurioidea) и морские лилии (Crinoidea) — широко распространены в морях и океанах нашей планеты (рис. 1). Морских звезд, морских ежей и голотурий легко встретить на приливно-отливной полосе всех полносоленых морей. В нашей стране иглокожих можно найти на литорали Баренцева и Белого морей, а также в дальневосточных морях. Эти животные очень чувствительны к солености, поэтому они не встречаются в пресных водах и в морях с пониженной соленостью, например в Балтийском и Черном.

Иглокожие распространены по всему Мировому океану, в том числе и на больших глубинах. Абиссальные равнины, простирающиеся на глубинах около 4 км и занимающие примерно 60% всей поверхности нашей планеты, тоже заселены этими животными. На мягких грунтах это голотурии из семейства Elpidiidae (рис. 1, к), а на твердых субстратах — стебельчатые морские лилии (рис. 1, м). Всего на нашей планете обитает не менее 6 тыс. видов Echinodermata.
И все же иглокожие — это одни из самых загадочных представителей животного царства. Начнем с того, что у большинства из них нет переднего и заднего концов тела. Обычная морская звезда — это хищник, который может двигаться в любую сторону, и каждый из ее лучей может в данный момент быть передним, а в любой другой момент — стать задним. У иглокожих нет ни брюшной, ни спинной стороны. Морские звезды и морские ежи обращены к субстрату ротовой стороной. Биологи называют ее оральной, а противоположную, на которой находится анус, — аборальной (рис. 2).

Рис. 2. Анатомия морской звезды: а — расположение мадрепорита в одном из интеррадиусов морской звезды и мадрепоровая плоскость животного; б — схема анатомической организации на срезе в мадрепоровой плоскости (красными стрелками указано преобладающее направление тока крови)
Иглокожие обладают радиальной симметрией. Она вторична — возникла в ходе сложнейших морфологических преобразований в процессе эволюции как надстройка над билатеральной симметрией. Ее слабые следы проявляются в расположении мадрепоровой пластинки, или мадрепорита (см. рис. 2, а) — небольшой пористой пластинки, которая находится в промежутке между двумя лучами морской звезды (биологи говорят — в интеррадиусе). Вполне возможно, что мадрепоровая плоскость иглокожих — это и есть остаток плоскости билатеральной симметрии далеких предков (см. рис. 2, а).
В анатомическом строении иглокожих и их физиологии множество загадок. Одна из них касается органов выделения у этих животных. В современных учебниках об этом либо ничего не сказано, либо лишь упомянуто, что у иглокожих имеются почки накопления — клетки, в которых продукты обмена накапливаются до конца жизни животного [1, 2]. Однако в процессе обмена веществ у них образуется много растворимых в воде соединений, которые надо постоянно выделять во внешнюю среду. Так, при распаде белков образуются азотистые вещества — аммиак или мочевина, которые растворяются в воде и вместе с мочой выводятся из организма наружу. У иглокожих продуктом азотистого обмена является аммиак — вещество очень токсичное, которое необходимо постоянно удалять из организма. Таким образом, почки иглокожим необходимы, и наша задача состояла в том, чтобы найти у них орган, который выполняет выделительную функцию.
Как функционируют выделительные органы беспозвоночных
Выделительные органы беспозвоночных тесно связаны с кровеносной системой — совокупностью полостей (гемоцелей) в толще соединительной ткани (рис. 3). Гемоцель отделена от других тканей слоем неклеточного вещества — базальной пластинкой, которая состоит из особого соединительнотканного белка — коллагена IV типа (см. рис. 3, а). Базальная пластинка — это стенка кровеносных сосудов. Снаружи на ней располагаются клетки мезодермального целомического эпителия, изнутри к ней могут прилегать клетки эндотелия (правда, у многих беспозвоночных эндотелиальная выстилка кровеносных сосудов отсутствует). Одевающие кровеносный сосуд снаружи клетки целомического эпителия имеют в цитоплазме сократимые волокна. Сокращение этих эпителиально-мышечных клеток приводит к возрастанию кровяного давления, и жидкость из кровеносных сосудов продавливается через базальную пластинку во вторичную полость тела — целом (см. рис. 3, а). Базальная пластинка — надежный фильтр, не пропускающий клетки крови, а также белковые молекулы, например гемоглобин и другие белки плазмы крови. Зато этот фильтр легко проницаем для мелких молекул, в том числе для аммиака.
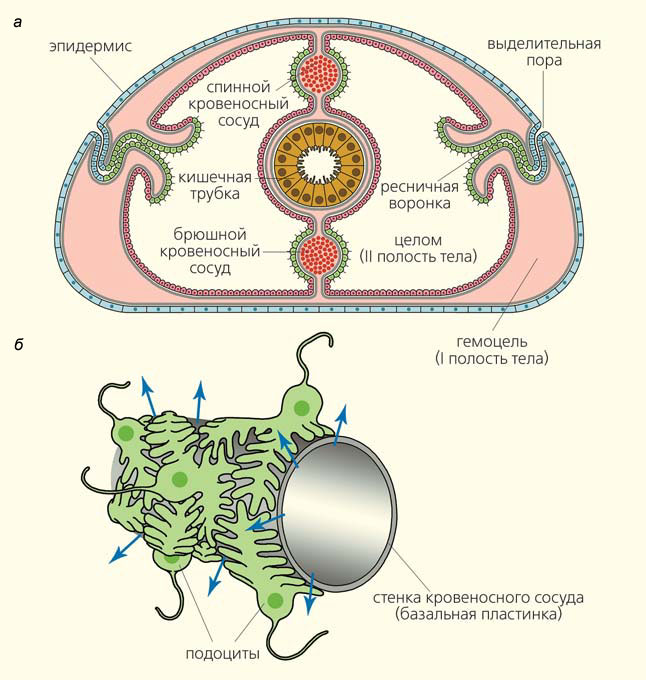
Рис. 3. Строение органов выделения у беспозвоночных: а — схема устройства и расположения мест фильтрации и выделительных воронок на поперечном срезе тела животного (серым цветом показаны базальные пластинки); б — строение кровеносного сосуда и подоцитов в выделительных органах, а также схема фильтрации (синие стрелки указывают направление фильтрации)
Обычно в кровеносной системе есть специальные места, предназначенные для такой фильтрации и связанные с органами выделения. В этих местах эпителиально-мышечные клетки целомической выстилки на поверхности кровеносных сосудов представлены особой разновидностью — клетками-подоцитами (см. рис. 3, б). От их основания отходят выросты, которые формируют многочисленные пальцевидные отростки. При этом отростки одной клетки входят в промежутки между отростками другой, приблизительно так, как сложенные в замок пальцы рук человека. Биологи так и называют это явление — интердигитация (от лат. inter — между и digitus — палец). Жидкость фильтруется из гемоцеля (т. е. из полости кровеносных сосудов) через базальную пластинку, проходит в узких промежутках между пальцевидными отростками подоцитов и попадает в целом (т. е. вторичную полость тела). Отростки подоцитов модифицируют проходящую жидкость: они извлекают из нее некоторые полезные вещества, а какие-то вредные соединения могут хранить в цитоплазме в виде крупных включений. Поэтому жидкость, попадающая в целом, в большей или меньшей степени отличается от плазмы крови. В зоологии беспозвоночных ту жидкость, которая прошла через базальные пластинки, принято называть первичной мочой, а ту, которая была модифицирована отростками подоцитов, — вторичной.
Собственно выделительные органы — это ресничные воронки, открывающиеся в целом (см. рис. 3, а). Биение ресничек загоняет целомическую жидкость в ресничные воронки, дальше она идет по длинному извитому каналу (где происходит значительная модификация мочи) и выводится во внешнюю среду.
Эта принципиальная схема приложима к большинству беспозвоночных, обладающих кровеносной системой и вторичной полостью тела — целомом. Так работают выделительные органы у кольчатых червей, почки моллюсков, щупальцевых и вторичноротых животных (Deuterostomia). Разумеется, в животном царстве много отклонений от этой схемы. Иначе устроены выделительные органы у животных, лишенных кровеносной системы (например, плоских червей), или у животных с редуцированным целомом (например, членистоногих), но мы не будем останавливаться на этих отклонениях.
Что служит почкой у иглокожих
Почка должна содержать несколько морфологических компонентов: во-первых, сеть кровеносных сосудов, во-вторых, целомическую полость, в которую фильтруется из них жидкость, и в-третьих, выделительный канал, который связывает целомическую полость с внешней средой. Оказывается, такой орган у иглокожих есть! Это так называемый осевой орган, связанный с мадрепоровой пластинкой (см. рис. 2, б). Располагается он между двумя лучами, т. е. в одном интеррадиусе, что придает анатомии иглокожих билатеральную симметрию (мы уже указывали, что, скорее всего, эта билатеральная симметрия унаследована радиально-симметричными иглокожими от их далеких билатерально-симметричных предков). Кровеносная сеть осевого органа представлена системой тянущихся от аборальной к оральной стороне тела гемоцельных пространств. Располагаются они между складками осевого целома. Бесцветная кровь иглокожих течет по этим гемоцельным пространствам преимущественно от аборальной к оральной стороне тела в околоротовое кровеносное кольцо (см. рис. 2, б). Направленное движение крови у иглокожих обеспечивается сокращениями сердца — пузырька, расположенного на аборальной стороне тела. Сердце принимает кровь из двух аборальных кровеносных колец — желудочного и генитального. Оно заключено в перикардиальный целом (или перикард), и эпителиально-мышечные целомические клетки на стенке сердца обеспечивают пульсацию этого органа. У иглокожих оно бьется нечасто — всего шесть раз в минуту [2]. Но этого достаточно, чтобы обеспечить направленное движение крови из двух аборальных кровеносных колец — желудочного и генитального — сквозь гемоцельные пространства осевого органа в околоротовое кровеносное кольцо. Оттуда кровь по радиальным сосудам течет в лучи, проходит сквозь кровеносную сеть, оплетающую печеночные выросты, и снова попадает в аборальные кровеносные кольца.
Гемоцельные пространства осевого органа отделены от осевого целома базальной пластинкой, на поверхности которой со стороны целома располагаются клетки целомической выстилки (рис. 4, а). В ее состав у иглокожих входит два типа клеток — подоциты и эпителиально-мышечные клетки (рис. 4, б и 5). Сокращения эпителиально-мышечных клеток обеспечивают то давление, благодаря которому жидкость из гемоцеля выдавливается в осевой целом. При этом жидкость проходит через фильтр, образованный базальной пластинкой, и модифицируется, проходя через узкие пространства между пальцевидными отростками подоцитов. Отметим, что отростки мышечных клеток могут пересекать гемоцельные пространства, соединяя складки целомического эпителия (разумеется, мышечные клетки всегда отделены от гемоцеля базальной пластинкой). Таким образом, мышечные клетки образуют единую пространственную сеть в пределах всего осевого органа (см. рис. 4, а).
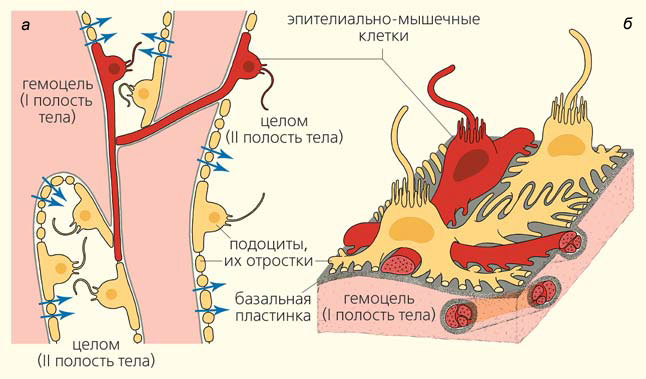
Рис. 4. Строение и функционирование осевого органа морской звезды Asterias amurensis: а — схема устройства целомической выстилки осевого органа на схематическом срезе; показаны отростки эпителиально-мышечных клеток, пронизывающие гемоцельные пространства, но отделенные от них базальной пластинкой (серый цвет); синими стрелками указано направление фильтрации; б — трехмерная схема тонкого строения целомического эпителия осевого органа [3]
Из осевого целома целомическая жидкость, содержащая продукты обмена, выводится во внешнюю среду через поры в мадрепоровой пластинке. Выделительная функция осевого целома доказана экспериментально: если ввести краситель в его полость, можно наблюдать, как краска выходит во внешнюю среду через мадрепоровую пластинку [4].
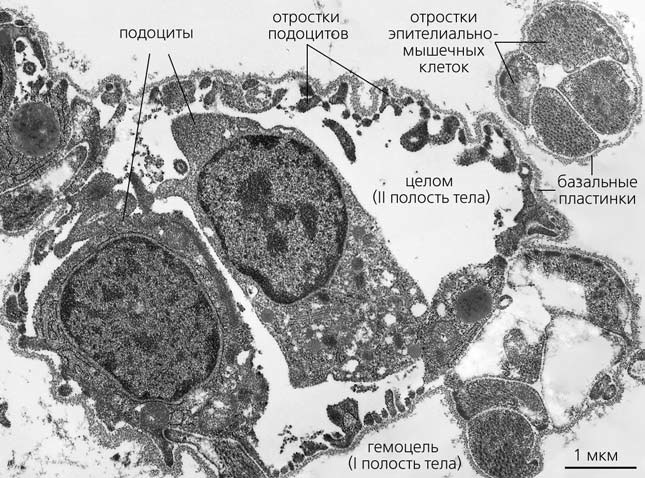
Рис. 5. Ультраструктурная фотография осевого органа морской звезды Asterias amurensis, сделанная с помощью трансмиссионного электронного микроскопа в межкафедральной лаборатории электронной микроскопии биологического факультета Московского государственного университета имени М. В. Ломоносова [3]. Фото Е. А. Егоровой
Как развивается почка иглокожих
Зоологи уверены, что иглокожие происходят от билатерально-симметричных предков, так как личинки иглокожих — билатерально-симметричные организмы (рис. 6, а). Вторичная радиальная симметрия формируется в результате сложных процессов при метаморфозе личинок. Они долго плавают в толще воды, питаясь одноклеточными планктонными водорослями.
Личинки иглокожих очень разнообразны по внешней форме, но имеют общие особенности. По их телу проходят так называемые ресничные шнуры. Они представляют собой ленту сложной конфигурации, состоящую из жгутиковых клеток, которые обеспечивают и локомоцию личинок, и их питание. Личинки иглокожих обладают обширным целомом, который связан с внешней средой каналом. Он открывается наружу отверстием — гидропором, расположенным на спинной стороне личинки (см. рис. 6, а).
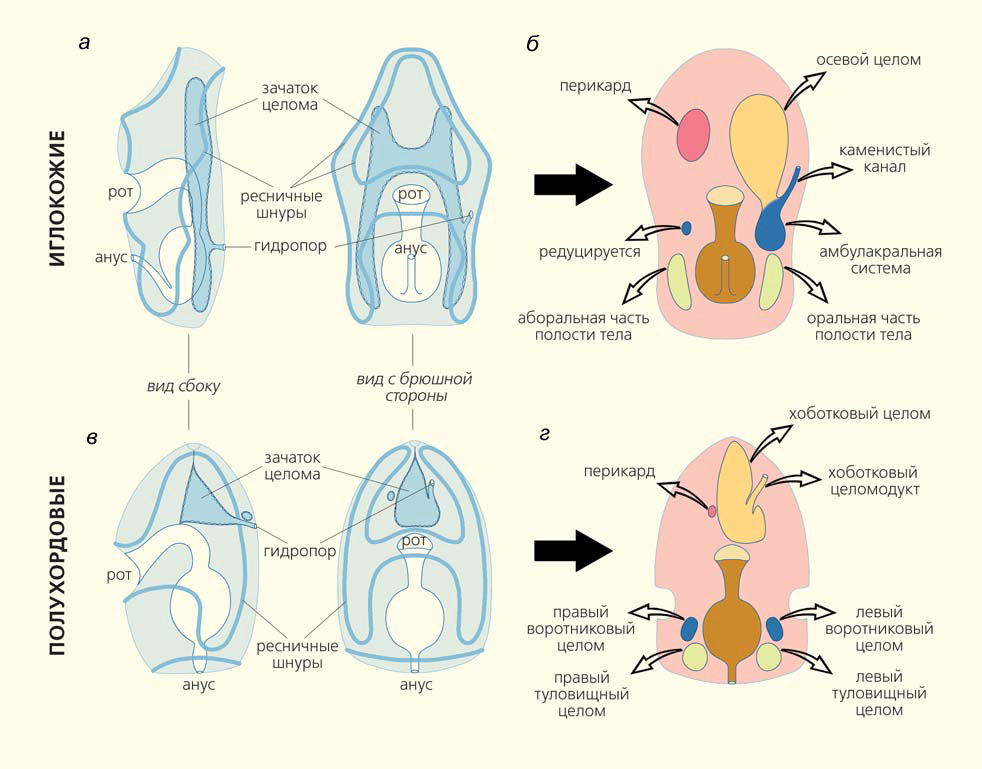
Рис. 6. Личиночное развитие иглокожих (Echinodermata) и полухордовых (Hemichordata): слева — личинка морской звезды (а) и кишечнодышащих (в); справа — отделы личиночного целома этих животных перед метаморфозом (б, г). Полыми стрелками показано, какие отделы целомов взрослых форм из них развиваются
Зачем нужен гидропор? Оказывается, это — выделительное отверстие. Жидкость из бластоцеля профильтровывается через базальную пластинку, попадает в целом, а затем выводится во внешнюю среду через гидропор. Нетрудно догадаться, что клетки целомической выстилки представлены подоцитами [5].
Перед метаморфозом целом расчленяется на три пары целомических мешков (см. рис. 6, б). Передние целомы личинки — это правый и левый аксоцели. В ее средней части располагаются правый и левый гидроцели, а в задней — лежат правый и левый соматоцели. Левый аксоцель и левый гидроцель всегда соединены между собой и связаны с внешней средой через гидропор. При метаморфозе (см. рис. 6, б) из левого аксоцеля развивается осевой целом, из левого гидроцеля — сложная водно-сосудистая (= амбулакральная) система иглокожих, а из правого аксоцеля — перикард. Правый гидроцель всегда редуцируется, никаких его производных у взрослых иглокожих нет. Что касается канала, соединяющего аксоцель и гидроцель личинки с внешней средой, то он дает начало характерному органу иглокожих — так называемому каменистому каналу. Самые задние целомы личинки — правый и левый соматоцели — дают начало полости тела, в которой помещается желудок, печеночные выросты и гонады взрослого животного.
Происхождение почки иглокожих
Повторим, что и внешнее строение, и анатомическая организация иглокожих очень необычны. Известный американский зоолог Э. Э. Рупперт писал о них так: «Иглокожие выглядят как инопланетяне, оказавшиеся на Земле, так удивительны их формы и функции» [2]. И все же по зоологической классификации иглокожие принадлежат к группе вторичноротых животных. Ближайшие родственники иглокожих — полухордовые (Hemichordata) и хордовые (Сhordata). Еще выдающийся русский биолог И. И. Мечников обнаружил, что личинки полухордовых (торнарии) и иглокожих удивительно сходны (см. рис. 6). Торнарии также долго плавают в толще воды и имеют ресничные шнуры сложной конфигурации (см. рис. 6, в). У личинок полухордовых тоже есть обширный целом, связанный с внешней средой каналом, который открывается наружу выделительной порой — гидропором. На основе сходства в строении личинок Мечников предложил объединить тип иглокожих и тип полухордовых в одну группу — амбулакралии (Ambulacralia) [6]. Современная молекулярная филогенетика полностью подтвердила предвидение Мечникова (рис. 7). И название Ambulacralia (в современных публикациях используется написание Ambulacraria) снова появилось на страницах зоологических журналов и учебников [7, 8].

Рис. 7. Схема филогении вторичноротых животных: типы иглокожих и полухордовых образуют группу амбулакралий
Перед метаморфозом целом личинки полухордовых приобретает расчленение, сходное с таковым у личинок иглокожих (см. рис. 6, г). В передней части личинки имеется обширный левый аксоцель, связанный с внешней средой гидропором, и маленький пузырек правого аксоцеля. В средней части личинки имеется пара целомов, которые можно сопоставить с гидроцелями, а в задней части личинки — пара целомов-соматоцелей.
Взрослые полухордовые совсем не похожи на иглокожих. В состав типа полухордовых входит два класса: кишечнодышащие и крыложаберные. Взрослые кишечнодышащие обитают в толще грунта и выглядят как червеобразные организмы, тело которых разделено на три отдела — хобот, воротник и туловище (рис. 8, а). А крыложаберные — это мелкие колониальные организмы, несущие на воротниковом отделе перистые щупальца (см. рис. 8, б). И у тех и у других имеется своеобразный рено-перикардиальный орган (лат. ren — почка, pericardium — околосердечная сумка). Авторы, пишущие на английском языке, называют этот орган heart-kidney — сердцепочка. В его состав входит хоботковый целом, который развивается из левого аксоцеля личинки полухордовых, перикард, формирующийся из правого аксоцеля, сердце и так называемый гломерулюс — сеть гемоцельных пространств между складками хоботкового целома (см. рис. 8, в). Сокращения сердца нагнетают кровь в сосуды гломерулюса. Его стенка образована базальной пластинкой и клетками целомической выстилки — подоцитами. Из гемоцельных пространств гломерулюса жидкость фильтруется через базальную пластинку, проходит между пальцевидными отростками подоцитов и попадает в хоботковый целом. Оттуда жидкость выводится во внешнюю среду через специальный канал (хоботковый целомодукт), который открывается наружу порой на спинной стороне животного.

Рис. 8. Полухордовые: а — Saccoglossus, представитель кишечнодышащих, б — Cephalodiscus, представитель крыложаберных; в — схема анатомической организации кишечнодышащих на срезе в сагиттальной плоскости (красными стрелками указано направление тока крови)
Рено-перикард полухордовых — несомненный гомолог осевого органа иглокожих. Он образуется из гомологичных целомов личинки и обладает существенным сходством в строении и функциях. Вероятно, предшественник рено-перикарда и осевого органа появился у общих предков иглокожих и полухордовых. Признаки, которые характеризуют родство двух групп животного царства, зоологи называют синапоморфиями. Можно считать, что рено-перикард, осевой орган, — это одна из важнейших синапоморфий группы Ambulacralia (= Ambulacraria), указывающая на глубокое филогенетическое родство полухордовых и иглокожих.
Итак, почка у иглокожих все-таки есть, — это давно известный осевой орган, а мадрепоровая пластинка — это выделительное отверстие иглокожих. Более того, оказалось, что почка иглокожих гомологична выделительному органу у представителей другого типа вторичноротых, а именно, у полухордовых. Таким образом, одна из загадок иглокожих разрешилась, но только одна... Иглокожие остаются, если использовать метафору Э. Э. Рупперта, «инопланетянами на морском дне», и их организация, развитие и эволюционная история таят еще множество тайн.
Образцы иглокожих, служившие материалом для исследований авторов, были собраны в Японском море при поддержке Российского научного фонда (соглашение № 14-50-00034), ультраструктурные исследования проведены при поддержке Российского фонда фундаментальных исследований (проекты 14-04-00366-a, 15-29-02601-офи-м).
Литература
1. Goldschmid A. Echinodermata // Spezielle Zoologie. Teil 1: Einzeller und Wirbellose Tiere / Eds W. Westheide, R. Rieger. Stuttgart, 1996. P. 778–834.
2. Ruppert E. E., Fox R. S., Barnes R. D. Invertebrate zoology. Belmont, 2004. V. 28. P. 872–929.
3. Ежова О. В., Егорова Е. А., Малахов В. В. Ультраструктурные доказательства выделительной функции осевого органа морских звезд (Asteroidea, Echinodermata) // ДАН. 2016. Т. 468. № 4. С. 470–473. DOI: 10.7868/S0869565216160271.
4. Cuénot L. Anatomie, éthologie et systématique des Échinodermes // Traité de Zoologie. Paris, 1948. V. 11. P. 3–272.
5. Balser E. J., Ruppert E. E., Jaeckle W. B. Ultrastructure of the coeloms of auricularia larvae (Holothuroidea, Echinodermata). Evidence for the presence of an axocoel // Biol. Bull. 1993. V. 185. P. 86–96.
6. Metschnikoff E. E. Über die Systematische Stellung von Balanoglossus // Zool. Anziger. 1869. Bd. 4. S. 139–143, 153–157.
7. Furlong R. F., Holland P. W. H. Bayesian phylogenetic analysis supports monophyly of Ambulacraria and of Cyclostomes // Zoological Science. 2002. V. 19. № 5. P. 593–599.
8. Nielsen C. Animal evolution: interrelationships of the living phyla. Oxford, 2001.











Рис. 1. Различные иглокожие: морские звезды — Asterias amurensis (а), Patiria pectinifera (б), Crossaster papposus (в); офиуры — Gorgonocephalus arcticus (г), Stegophiura nodosa (д), Ophiothrix savignyi (е); морские ежи — Strongylocentrotus pallidus (ж), S. purpuratus (з), Heterocentrotus mamillatus (и); голотурии — Scotoplanes sp., или «морская свинья» (к), Apostichopus japonicus (л); морские лилии — Neocrinus decorus (м), Heterometra savignyi (н). Фото А. А. Семенова, кроме и (noaa.gov), к (tumblr.com), м (photoflowery.ru)