Когда, где и зачем поют самки птиц
Ирина Бёме, Мария Горецкая
«Природа» №7, 2015
 Ирина Рюриковна Бёме — доктор биологических наук, профессор кафедры зоологии позвоночных Московского государственного университета им. М. В. Ломоносова. Область научных интересов — биокоммуникация, систематика и филогения птиц. |
 Мария Яковлевна Горецкая — кандидат биологических наук, научный сотрудник Звенигородской биостанции Московского государственного университета им. М. В. Ломоносова. Орнитолог, специалист в области биокоммуникации, экологии и поведения птиц. |
Вряд ли найдется человек, который никогда не слышал пения птиц. Приход весны возвещается их бодрым хором, звучащим как в лесу, так и в городской среде. Большинство поющих птиц относятся к воробьинообразным (Passeriformes) — одному из широко распространенных и высокоорганизованных отрядов в классе птиц.

Традиционно вокализацию воробьинообразных подразделяют на позывы — короткие, простые звуки, издаваемые в самых разнообразных ситуациях в течение всего года, — и песню, которая длится дольше, имеет сложную структуру и связана в основном с репродуктивным периодом в жизни птиц. Песня выполняет разные, иногда противоречивые функции [1]. Одна из наиболее важных и очевидных — это обеспечение встречи половых партнеров, привлечение самки и, возможно, образование пары [2–4]. У многих видов самцы продолжают петь, когда пара уже создана. Предполагается, что в этом случае песни адресованы соседям и связаны с демонстрацией занятости территории [5]. Считается, что видоспецифическая структура песни необходима для обеспечения репродуктивной изоляции близких видов птиц [4], причем для формирования такой песни важно, чтобы птенцы слышали в чувствительный период пение самцов своего вида [4, 6].
Многофункциональность песен, видимо, приводит к усложнению их структуры и повышению индивидуальной изменчивости их компонентов [1]. Происхождение песни связывают с половым поведением самцов, а именно — привлечением самки [7]. Предполагали, что эволюция песни шла от позывов самцов к сложной территориальной песне, при этом в ее структуру вовлекались различные позывы, которые постепенно заменялись в репертуаре на другие сигналы [8].
Исторически работы по изучению песни проводились в Европе, где у большинства видов поют только самцы, поэтому ее роль обсуждалась только применительно к ним. Однако даже в умеренном поясе поющие самки — не такая уж редкость [9], а уж в тропиках — скорее норма, чем исключение из правил [10–13]. В прошлом году в Nature Communications вышла статья, в которой авторы на основе анализа песен воробьинообразных птиц утверждают, что пение самок — древний признак для этого отряда [14].
Мы проанализировали случаи пения самок у 1309 видов, относящихся к двум основным группам отряда воробьинообразных (16 семействам подотряда певчих воробьиных и пяти — кричащих), обитающих на разных материках. Сведения о них мы почерпнули из описаний в сводке HandBook of the Birds of the World [15]. Цель нашего исследования — понять эволюционные предпосылки этого феномена и его связь с особенностями биологии видов и путями распространения основных групп воробьиных. Семейства были выбраны так, чтобы в разных частях света число изученных видов было сходным.
Как и зачем поют самки
Не только у воробьинообразных поют самки. Известны случаи их брачной вокализации вплоть до смены половых ролей и у других птиц (плавунчиков, трехперстков, тинаму и др.), когда самки первыми прилетают в район гнездования и громкой песней привлекают самцов [16]. Иногда в брачный период птицы поют дуэтом, причем удивительно согласованно, хотя координируют свои партии по-разному. У некоторых видов самка и самец издают звуки, точно чередуя их во времени, а кажется, будто звучит голос одной птицы. Такой дуэт называют антифональным. У рыбного филина (Ketupa blakistoni), крупнейшей совы Дальнего Востока, партию начинает самец низким криком, ему вторит самка, затем снова слышится уханье самца, и сразу же протяжно гудит самка. Таким образом, вся песня состоит из четырех сигналов, из которых первый и третий принадлежат самцу, а второй и четвертый — самке [17]. У других видов партнеры не столь четко координируют свои партии, и звуки участников могут накладываться друг на друга.

Эфиопский певчий сорокопут. Самцы и самки этого вида почти неотличимы
У воробьинообразных также отмечены случаи дуэтного пения. Впервые оно было описано у нескольких видов африканских сорокопутов (Laniarius). Так, согласованный дуэт исполняют супруги эфиопского певчего сорокопута (L. aethiopicus). Однако самки воробьинообразных поют и соло. Дуэты встречаются только у 52% видов с поющими самками, что составляет 12% от всех проанализированных нами видов отряда. Дуэты в большей степени характерны для мономорфных видов (60%), в меньшей — для видов с половым диморфизмом (41%). Дуэтом поют чаще пары оседлых видов (58%), чем мигрирующих (17%).
Самки могут петь не только для поддержания контакта в паре и синхронизации брачной активности [18, 19], но и для привлечения самцов [11, 12, 20, 21], для приглашения их к копуляции [12, 22], а также для защиты территории [22]. Например, у прекрасного расписного малюра (Malurus cyaneus) в ответ на проигрывание видоспецифической песни на их территории самки ведут себя даже агрессивнее самцов [22, наши данные]. Они подлетают к громкоговорителю и пытаются его атаковать, а также вступают в песенную дуэль с громкоговорителем — исполняют свою песню вслед за проигранной.

Чета прекрасных расписных малюров
Распространенность
Даже в тропических широтах есть семейства птиц, в которых поют самки всех видов, и такие, в которых это бывает довольно редко (5–25% видов). В южных широтах обоих полушарий рекордсмены по пению — самки семейств африканских кустарниковых сорокопутов (73% видов), американских трупиалов (57%) и кардиналов (42%). В Австралии, где на первый взгляд поют самки практически всех видов, их недостаточная изученность не позволяет провести точную оценку встречаемости поющих самок. Это в том числе может быть связано с отсутствием полового диморфизма у многих видов. Если не брать во внимание малоизученные виды, наиболее часто (67% видов) поют самки австралийских славок (Maluridae) и у всех птиц-лир (Menuridae). Среди проанализированных нами евразийских видов самки поют значительно реже: у 26% видов подсемейства чекановых (Saxicolinae), по 3% — славковых (Sylviidae) и синиц (Paridae). Например у многочисленной в Европе зарянки (Erithacus rubecula), мономорфного вида, поют и самцы, и самки.

Красный кардинал. Песня самца (слева) отдаленно напоминает соловьиную. Самки тоже поют, и их песня также довольно разнообразна.
В целом, по нашей оценке, доля видов в отряде воробьинообразных, у которых отмечены поющие самки, не превышает 30%. Однако несомненно, что этого вполне достаточно, чтобы разрушить устоявшиеся представления о пении как о прерогативе самцов.
Мировое распространение поющих самок позволяет выдвинуть гипотезу об эволюции их пения у воробьинообразных птиц. По мнению наших зарубежных коллег, становление и распространение кричащих и певчих птиц отряда воробьинообразных шло после распада Гондваны в разное время и разными путями [23]. Кричащие птицы вначале заселили Африку и Азию и лишь затем Южную и Северную Америку. В Евразии мы нашли 11% таксонов с поющими самками, в то время как в Америке — 14%. Разные ветви певчих птиц сначала освоили Австралию, потом Евразию, затем Африку и наконец Северную и Южную Америки [24]. Наш анализ показал, что в Австралии доля самок-певуний составляет 28%, менее всего в Евразии (10%), а максимальное количество в Африке (33%) и Америке (37%). Таким образом, самки-вокалисты чаще встречаются в более молодых группах как кричащих, так и певчих птиц.
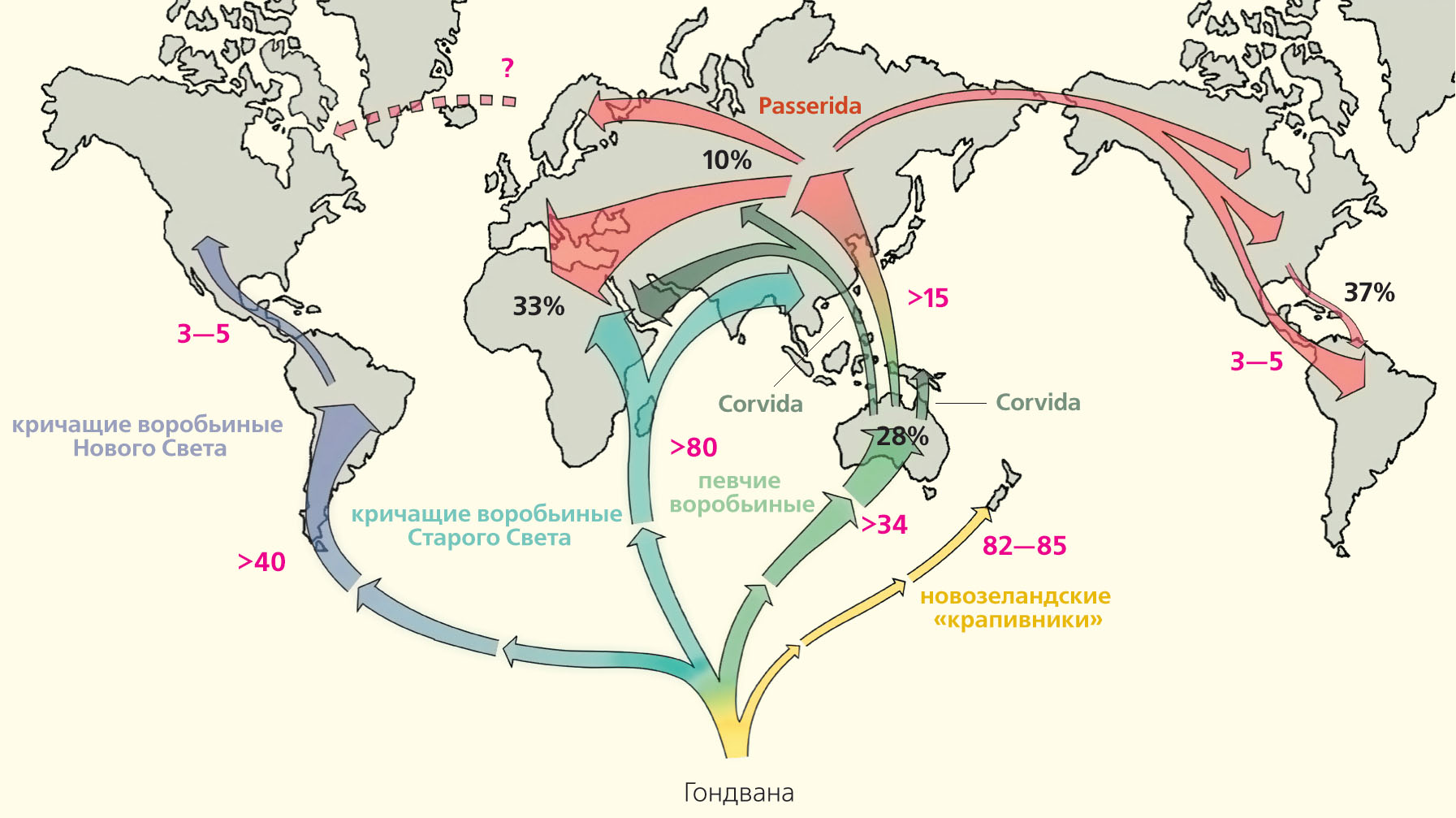
Предполагаемые пути расселения основных групп птиц отряда воробьинообразных после распада Гондваны. Схема создана на основе данных филогенетических отношений и биогеографии (розовым цветом указано время расхождения ветвей в млн лет назад; Passerida и Corvida — группы певчих воробьиных) [23]. Кричащие птицы вначале заселили Африку и Азию и лишь затем Америку, а певчие воробьиные начали свой путь в Австралии. Там, по нашим подсчетам, сейчас обитает около 28% видов с поющими самками, в Евразии — 10%, Африке — 33%, Америке — 37% [24]
Когда выгодно петь самкам
По нашим данным, чаще поют самки оседлых видов (83% у певчих птиц, 100% у кричащих), реже — у перелетных (17% у певчих), что неудивительно. Птицы умеренной зоны с приближением холодного времени года улетают на юг, а с наступлением весны возвращаются на места гнездования. У многих воробьинообразных самцы прилетают раньше, занимают территории и охраняют их. Самки присоединяются к ним чуть позже и выбирают партнера, оценивая не только его «личные качества», но и территорию, которую он сумел занять. Супружеская пара у таких видов каждый год формируется заново. Время на размножение у перелетных видов очень ограничено, за два-три месяца птицы должны успеть выбрать территорию, создать пару, выкормить потомство и перелинять. Видимо, в такой ситуации отбор идет против пения самок.

Большая птица-лира, или обыкновенный лирохвост
Пернатые обитатели тропиков, наоборот, могут весь год держаться на одной территории, которую должны постоянно охранять, что проще делать вдвоем, чем в одиночку, и в такой ситуации пение самок становится выгодным. Пример тому — североамериканский домовой крапивник (Troglodytes aedon), самки и самцы которого внешне неразличимы. На юге ареала (в Центральной Америке) эти птицы, во-первых, оседлы, во-вторых, моногамны. Супруги вместе защищают территорию круглый год, для этой цели им служит отлаженный, скоординированный дуэт. А вот в северных популяциях самцы полигамны, они прилетают на места гнездования, занимают и охраняют каждый год новый участок. В этих популяциях самки крапивника не поют.
По данным исследователей, анализировавших евразийские виды, пение самок связано с наличием полового диморфизма в окраске, причем только в тех случаях, когда она обусловлена липохромовыми пигментами, которые обеспечивают желтый, красный и рыжий цвета [1]. Формирование такой окраски связано с содержанием каротиноидов в рационе птиц, и ее вариации, возникающие в процессе полового отбора, могут свидетельствовать о средовых влияниях и сигнализировать о физиологическом состоянии особи. Наличие липохромов в окраске, по мнению авторов статьи, может быть «честным» признаком качества ее носителя. Нам не удалось выявить связь между пением самок и наличием липохромов в окраске, однако обнаружена корреляция между половым диморфизмом и пением самок. Возможно, результаты, полученные нашими зарубежными коллегами, также связаны не с окраской как таковой, а с выраженностью полового диморфизма.
Наши данные не поддерживают гипотезу о том, что пение самок — древний признак для всех воробьинообразных птиц. Видимо, оно возникало независимо в разных семействах, причем для каких-то из них (например, семейства трупиаловых [25, 26]) действительно может быть древним.

Самец красноплечего трупиала (слева) в характерной позе — с раскрытым веером хвостом. Самец ярче (особенно заметны желтые и красные отметины на плечах и крыльях) и крупнее самки. Она хоть и невзрачна на вид по сравнению с партнером (похожа на крупного воробья), зато не уступает ему в пении
Изначально акустические сигналы характерны для обоих полов в классе птиц, во многих отрядах распространены вокальные дуэты*. В отряде воробьинообразных пение получило наибольшее распространение и очевидно, что отбор на воспроизведение и восприятие песни шел не только среди самцов, но и среди самок. Известно, что у самок в процессе онтогенеза также развиваются вокальные центры головного мозга, а при стимуляции тестостероном в норме непоющие самки начинают петь [4]. Понятно, что способности к пению должны развиваться у самок и самцов параллельно, а проявление этого признака зависит от конкретной ситуации. Самки кричащих воробьиных после проникновения в Старый Свет практически утрачивают способность к пению (надо заметить, что песни и самцов этих видов крайне примитивны и просты), в то время как в Америке доля поющих самок увеличивается. Для певчих воробьиных наблюдается сходная картина. Исходя из вышесказанного, гипотеза о древности признака пения самок применительно к отряду воробьинообразных выглядит несколько странно. Видимо, пение самок независимо проявляется в различных семействах птиц этого отряда, как кричащих, так и певчих.
Авторы благодарны сотрудникам Университета Монаш (Австралия) Александре Павловой и Полу Санок и Университета Дикон (Австралия) Кейт Бучанон за предоставленную возможность работать в составе их экспедиции в штате Виктория в Австралии.
Работа выполнена при поддержке Российского фонда фундаментальных исследований (проект 14304300108а).
* Подробнее см.: Брагина Е. В., Бёме И. Р. Дуэты у животных — почему птицы поют хором? // Природа. 2010. № 6. С. 13–18.
Литература
1. Ильичев В. Д. Биоакустика птиц. М., 1972.
2. Мальчевский А. С. К вопросу о голосовой имитации у птиц // Сложные формы поведения. М.; Л., 1965. С. 139–143.
3. Тинберген Н. Социальное поведение животных. М., 1993.
4. Catchpole C. K., Slater P. J. B. Bird song: biological themes and variations. Cambridge, 2008.
5. Thorpe W. H. Bird-song: the biology of vocal communication and expression in birds. L.; NY, 1961.
6. Бёме И. Р., Горецкая М. Я. Песни птиц: Учебное пособие. М., 2013.
7. Дарвин Ч. Происхождение человека и половой отбор. М.; Л., 1951. Т. 5. С. 199–658.
8. Симкин Г. Н. Актуальные проблемы изучения звукового общения птиц // Орнитология. 1982. Вып. 17. С. 111–129.
9. Garamszegi L. Z., Pavlova D. Z., Eens M., Møller A. P. The evolution of song in female birds in Europe // Behav. Ecol. 2007. V. 18. P. 86–96. doi: 10.1093/beheco/arl047.
10. Kroodsma D. E., Viellard J. M. E., Stiles F. G. Study of bird song in the Neotropics: urgency and opportunity // Ecology and evolution of acoustic communication in birds / Eds D. E. Kroodsma, E. H. Miller. Ithaca, 1996. P. 269–281.
11. Morton E. S. A comparison of vocal behavior among tropical and temperate passerine birds // Ecology and evolution of acoustic communication in birds / Eds D. E. Kroodsma, E. H. Miller. Ithaca, 1996. P. 258–268.
12. Langmore N. E. Functions of duet and solo songs of female birds // Trends Ecol. Evol. 1998. V. 13. P. 136–140. doi: 10.1016/S0169-5347(97)01241-X.
13. Slater P. J. B., Mann N. I. Why do the females of many bird species sing in the tropics? // J. Avian Biol. 2004. V. 35. P. 289–294. doi: 10.1111/j.0908-8857.2004.03392.x.
14. Odom К. J., Hall M. L., Riebel K. et al. Female song is widespread and ancestral in songbirds // Nature Communications. 2014. V. 5. № 3379. doi: 10.1038/ncomms4379.
15. HandBook of the Birds of the World / Eds J. del Hoyo, A. Elliott, D. Christie. Barcelona, 2004–2011. V. 8–12, 16.
16. Ильичев В. Д., Карташев Н. Н., Шилов И. А. Общая орнитология. М., 1982.
17. Пукинский Ю. Б. Жизнь сов. Л., 1977.
18. Ratcliffe L., Otter K. Sex differences in song recognition. Vocal recognition of neighbors by territorial passerines // Ecology and Evolution of Acoustic Communication in Birds / Eds D. E. Kroodsma, E. H. Miller. NY, 1996. P. 339–355.
19. Silva H. G. de, Marantz C. A., Perez-Villafana M. Song in female Hylorchilus wrens // Wilson Bull. 2004. V. 116. P. 186–188.
20. Levin R. N. Song behaviour and reproductive strategies in a duetting wren, Thryothorus nigricapillus: II. Playback experiments // Anim. Behav. 1996. V. 52. P. 1107–1117.
21. Eens M., Pinxten R. Female song for mate attraction: an overlooked phenomenon? // Trends Ecol. Evol. 1998. V. 13. P. 322–323.
22. Cooney R., Cockburn A. Territorial defence is the major function of female song in the superb fairy-wren (Malurus cyaneus) // Anim. Behav. 1995. V. 49. P. 1635–1647.
23. Ericson P. G. P., Christidis L., Cooper A. et al. A Gondwanan origin of passerine birds supported by DNA sequencies of the endemic New Zealand wrens // Proc. R. Soc. Lond. B. 2002. V. 269. P. 235–241. doi: 10.1098/rspb.2001.1877.
24. Бёме И. Р., Горецкая М. Я. Пение самок воробьинообразных птиц: анцестральность или эпифеномен пения самцов? // Журн. общей биологии [в печати].
25. Price J. J. Evolution and life-history correlates of female song in the New World blackbirds // Behav. Ecol. 2009. V. 20. P. 967–977.
26. Price J. J., Lanyon S. M., Omland K. E. Losses of female song with changes from tropical to temperate breeding in the New World blackbirds // Proc. R. Soc. Lond. B.Biol. Sci. 2009. V. 276. P. 1971–1980.

Пение птиц
-
 13.12.2020Пение самок птиц, или почему поют только самцыМария Монахова • Видеотека
13.12.2020Пение самок птиц, или почему поют только самцыМария Монахова • Видеотека -
 20.03.2019Птицы осваивают пение, наблюдая за реакцией сородичейАнтон Морковин • Новости науки
20.03.2019Птицы осваивают пение, наблюдая за реакцией сородичейАнтон Морковин • Новости науки
-
 06.12.2018Самцам птиц приходится выбирать между привлекательной окраской и красотой песниАнтон Морковин • Новости науки
06.12.2018Самцам птиц приходится выбирать между привлекательной окраской и красотой песниАнтон Морковин • Новости науки
-
 01.11.2018Алгебра и гармония в песне восточного соловьяВладимир Иваницкий, Ирина Марова, Владислав Антипов • Библиотека • «Природа» №2, 2014
01.11.2018Алгебра и гармония в песне восточного соловьяВладимир Иваницкий, Ирина Марова, Владислав Антипов • Библиотека • «Природа» №2, 2014
-
 23.10.2018Зебровым амадинам для успешного обучения пению необходимо видеть учителяАлександр Марков • Новости науки
23.10.2018Зебровым амадинам для успешного обучения пению необходимо видеть учителяАлександр Марков • Новости науки
-
 25.01.2018Когда, где и зачем поют самки птицИрина Бёме, Мария Горецкая • Библиотека • «Природа» №7, 2015
25.01.2018Когда, где и зачем поют самки птицИрина Бёме, Мария Горецкая • Библиотека • «Природа» №7, 2015
-
 15.11.2017Воркование голубяАлексей Опаев • Картинки дня
15.11.2017Воркование голубяАлексей Опаев • Картинки дня
-
 18.08.2017Самцы зебровых амадин слушают других самцов, а самки — только супругаТатьяна Романовская • Новости науки
18.08.2017Самцы зебровых амадин слушают других самцов, а самки — только супругаТатьяна Романовская • Новости науки
-
 21.02.2017В пении флейтовых птиц обнаружены музыкальные принципыАнтон Морковин • Новости науки
21.02.2017В пении флейтовых птиц обнаружены музыкальные принципыАнтон Морковин • Новости науки
-
 13.12.2016В птичьей песенке наследуются интервалы, а сами звуки выучиваютсяЕлена Наймарк • Новости науки
13.12.2016В птичьей песенке наследуются интервалы, а сами звуки выучиваютсяЕлена Наймарк • Новости науки







Поющая зарянка. Здесь и далее использованы фотографии Викисклада