Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Как появились тилакоиды цианобактерий

Появление тилакоида - важного морфологического элемента многократно увеличило фотосинтетическую мощность цианобактерий, игравших основную роль в переходе от темного фотосинтеза к светлому и образовании кислородной атмосферы и растительного покрова Земли. В предыдущих работах автора было показано, что фрактальные геометрические свойства точечных множеств могут определять характер морфологической эволюции биологических организмов. Для эволюции цианобактерий таким свойством явилось уменьшение фрактального показателя множеств при групповом размещении растущего числа точек множества и постоянном числе их в группе. В этом случае виртуальный общий размер фотосинтетического аппарата цианобактерий должен стать во много раз больше, чем ограниченный размер самой бактерии. Невозможность такого экстенсивного роста побудило цианобактерии к эволюционным изменениям своей внутренней морфологии – созданию тилакоидов с дополнительными площадями для размещения новых фотосинтезирующих “точек”. Обсуждаются новые опубликованные экспериментальные данные, подтверждающие справедливость этого вывода.
Читателю, решившему просмотреть список ссылок на статьи автора к этой статье про цианобактерии и их тилакоиды, может показаться странным, что история начинается с довольно простой 3D секционной модели размещения в пространстве зеленой биомассы реальной ели. Что роднит эти визуально разные элементарные фотосинтезирующие объекты (“точки” [0,1) – мономеры димеров PSII и тримеров PSI, отрезки линий (1,2) – хвоя, плоские кусочки [2,3) - листья) на всем протяжении их эволюции? Математические свойства размещений их систем зеленых фотосинтезирующих элементов в доступном в нашем трехмерном мире пространстве описываются фрактальной геометрией на всем интервале [0,3] изменения фрактального параметра μ, который в случае [0,1) связывает число точек Np с размером H множества точек: Np ̴̴ Hμ.
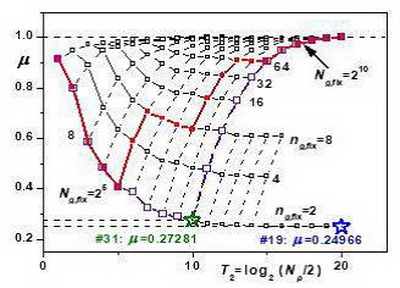
В статье ЖОБ, 2016 (Сколько веток у ели?; bioRxiv, 2018) было продемонстрировано влияние размещения множества точек группами на характер зависимости μ(Np) (где Np=Ng*ng, Ng – число групп, ng – число точек в группе) при увеличении Np, (рис. 1). Если увеличение Np достигается увеличением Ng при фиксированном числе точек в группе ng,fix, то μ(Np) уменьшается, стремясь к μ2 ≈ 0.25 при ng,fix=2 и к μ3 ≈ 0.37 при ng,fix=3. Таким образом при увеличении числа точек Np в два раза размер множества должен увеличиться в 16 и 9 раз соответственно. Поскольку размер цианобактерии ограничен, простое увеличение Np фактически невозможно.
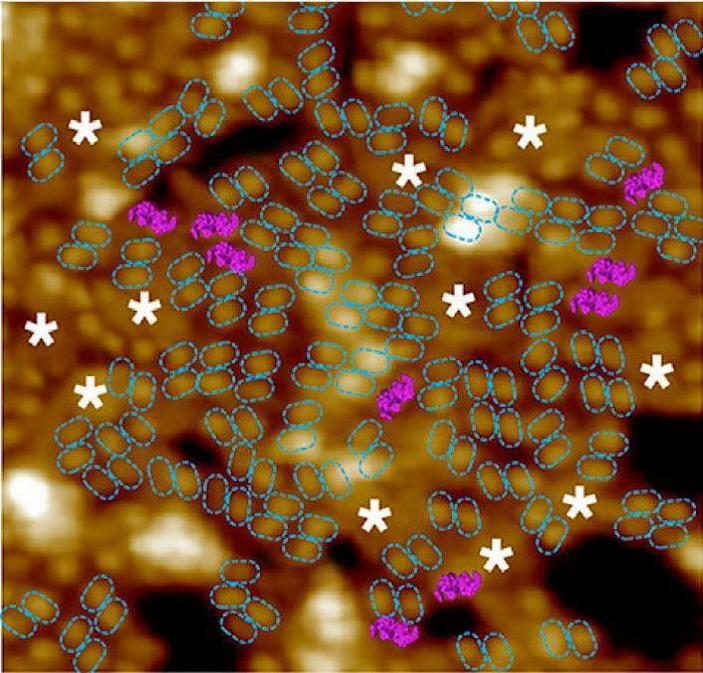
Путь к эволюции фотосинтеза был открыт с созданием нового морфологического элемента цианобактерии – тилакоида: впячивая участок поверхности цитоплазматической мембраны и искривляя её внутреннюю поверхность, предки цианобактерий начали создавать дополнительные поверхности для размещения фотосинтезирующих элементов – тилакоидов. Для этого бактериями был создан специальный белок CURT1, недавно найденный и у растений и это есть косвенное подтверждение модельных заключений. У цианобактерий (и у растений) обнаружено групповое размещение “точек”-мономеров в виде димеров и тримеров (рис. 2). С использованием атомно-силовой микроскопии AFM путем буквального ощупывания мембраны получены изображения размещений димеров и тримеров на мембране группами для нескольких видов бактерий и это уже прямое подтверждение модели, Casella, S. et al. Mol. Plant. 10. 2017. Более того, в другой работе (MacGregor-Chatwin, C. et al. Plant Cell. 29. 2017) тем же AFM-методом было получено, что для другого вида бактерий числа тримеров и димеров, размещенных на мембране соотносятся как N3:N2 = 4. С использованием значений µ3 и µ2 из рис. 1 величина этого соотношения вычислена как 4.008 и при этом показано, что тримеры и димеры расположены на одной площадке цитоплазматической мембраны, хотя и на противоположных её сторонах.
Популярные синопсисы