Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Растения, сбежавшие с земли

Статья представляет собой первый на русском языке обширный обзор и обобщение современных взглядов на феномен эпифитизма сосудистых растений. В нем, на основе задействованного очень широкого круга литературы (более 280 источников), обсуждается проблема происхождения эпифитизма, его экологических границ и современного понимания экоморфологических особенностей эпифитов. Рассмотрены различные подходы к классификации эпифитов и механизмы существования растений, порвавших с наземными источниками минерального питания и воды.
Побег с земли мечтает совершить не только Илон Маск. Намного ранее (десятки, сотни миллионов лет назад, точно не известно) это совершили многие растения. Правда сбежали они не на Марс, а всего лишь порвав «родовую пуповину» всех растений – связь с землей и переселившись в кроны деревьев. Это создало интересный пример своего рода «сферического коня в вакууме» среди растений, когда все органы растительного организма находятся в воздушной среде, и он почти полностью изолирован от потока минерального питания и воды, поступающих из почвы.
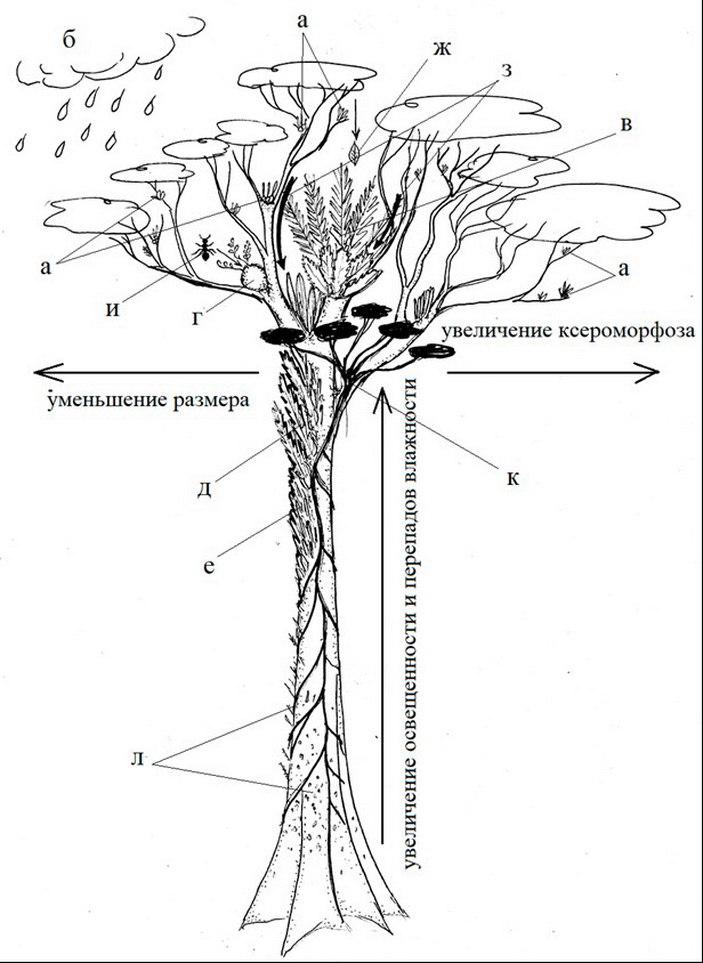
Освоив новые высоты, в которых они почти не встретили конкуренции, эпифиты неожиданно дали непропорционально большой вклад в большинстве глобальных центров биоразнообразия растений (Taylor et al., 2021). До недавнего времени насчитывалось около 27600 видов сосудистых эпифитов из 73 семейств и 913 родов, что составляет 9% от всех видов сосудистых растений (Zotz, 2013). Однако публикация первого полного списка сосудистых эпифитов показало существенно большее их число: 28227 цветковых и 3084 споровых видов (Zotz, 2021). Основная часть сосудистых эпифитов приходится на папоротники и однодольные, особенно на семейства Orchidaceae, Bromeliaceae, Araceae (Zotz, 2013). Небольшое число эпифитов и полуэпифитов приходится на двудольные растения, в том числе, из семейств Ericaceae, Gesneriaceae, Piperaceae (Krömer, Gradstein, 2016). Узкие экологические рамки в ходе эволюции привели и к реализации ряда биоморфологических схем (Рис. 1), связанных с разного типа «ответами» на вызовы дефицита влаги и питательных веществ (Рис. 2).
Традиционное пренебрежение сосудистыми эпифитами в отечественной научной литературе, можно объяснить и тем фактом, что они почти полностью отсутствуют на территории исторической России (включая РИ и СССР). Большая часть биоразнообразия эпифитов сосредоточена в тропических и субтропических областях Земли (Benzing, 1990; Zotz, 2016). При этом мало изучены их экосистемная и эдификаторная роли. Вокруг эпифитов происходит депонирование впечатляющего массива органического вещества в кронах леса. Нами высказана мысль, что подвешенные почвы выполняет ту же функциональную роль, что и лесная подстилка в лесах более высоких широт и которая в тропических лесах отсутствует (Eskov et al., 2021). Таким образом, эпифитные сообщества (да и в целом подвешенная биота) являются не только существенным источникам биоразнообразия тропических лесов, но и недооценённым депозитарием органического вещества, структурированного эпифитами. Вокруг накопленного органического вещества возникают многочисленные трофические и консортивные связи с участием насекомых, грибов, позвоночных, низших растений. Наконец, многочисленные организмы филлосферы (печеночники, мхи, лишайники, бактерии, грибы и другие микроорганизмы), которые мы на прямую не включаем в эпифитное сообщество, но которые можно отнести вместе с эпифитами к подвешенной биоте, оказывают существенное влияние на фотосинтетическую стратегию, гидравлику, поглощение ресурсов, температуру листа, защитные и многие другие свойства (Jones, Dangl, 2006; Lambers et al., 2008; Friesen et al., 2011; Sawinski et al., 2013; Kembel et al., 2014 и др.). Эпифиты и их консортивный круг являются ярким и недооценённым примером «темного биоразнообразия» (dark biodiversity) для тропических экосистем и сулят нам много сюрпризов.
Перспективы и актуальность изучения эпифитов многоразличны. С одной стороны, почти отсутствует цельная палеоботаническая картина происхождения эпифитов, что ставит вопрос о причинах происхождения их впечатляющего разнообразия. Эпифиты играют важную роль в создании глобального градиента широтного разнообразия растений (Taylor et al., 2021). Это делает их крайне уязвимой группой, ведь, будучи структурно зависимыми растениями, они целиком и полностью зависят от лесов произрастания. А 60% тропических лесов еще 20 лет назад были признаны деградировавшими (ITTO, 2002). Судя по многочисленным исследованиям, эпифиты служат хорошим диагностическим критерием состояния лесов, как наиболее уязвимая экогруппа (например, Leao et al., 2014). На примере семейства бромелиевых было показано, какую огромную роль в экосистемных услугах (ecosystem services) могут играть эпифиты (Ladino et al., 2019). Кроме того, в связи с изменением климата и увеличением антропогенных выбросов, непонятно, как это скажется на эпифитах. Предполагалось, что возможно повышение содержания СО2 сделает более эффективным механизмы водопользования у эпифитов (иными словами увеличивает их ксерофильность). Но в целом однозначного доказательства о положительном влиянии повышенного содержания СО2 на эпифиты найдено не было (Monteiro et al., 2009; Raveh et al., 1995; Li et al., 2002; Zotz et al., 2010).

Привлечение внимания к тропическим экосистемам очень актуально для отечественной науки. Не смотря на традиционно высокий уровень биологических дисциплин в России, в нашей стране почти отсутствует школа структурных исследований тропических экосистем. Это выражается например в том, что в рамках традиционного отечественного биоморфологического подхода многие современные взгляды на тропические растения можно понять только контекстуально (Рис. 3). В попытке отчасти изменить этот тренд и написана обсуждаемая обзорная статья. Она преследует несколько целей. Во-первых, познакомить русскоязычных ботаников с состоянием науки вокруг изучения сосудистых эпифитов, ведь традиционно все, что касается тропических экосистем, остаётся по большей части вне поля внимания отечественной науки. Во-вторых, мы постарались подчеркнуть и выделить те существенные черты (от физиологических до популяционных), которые отличают сосудистые эпифиты от наших северных растений. В-третьих, мы ставили своей целью систематизировать многочисленную литературу о сосудистых эпифитах вокруг определенной эволюционной схемы, в рамках которой эпифиты стали совершенно уникальным биологическим объектом, когда впечатляющее биоразнообразие было достигнуто в очень узких экологических рамках. Эти рамки могут быть сведены к трем экологическим «вызовам» обитания на стволе дерева: (1) баланс факторов увлажнения и освещенности, (2) строго лимитированное минеральное питание, (3) закрепление растений и их диаспор в кронах деревьев.
Все эпифиты для освоения эпифитных местообитаний должны были решить эти проблемы, преобразуя не только свой габитус, но и систему воспроизведения и прорастания (Рис. 2). Таким образом, структура данной работы идет вслед за этой схемой: мы рассматриваем сначала сам феномен эпифитизма (включая проблему классификации эпифитов), переходим к их экофизиологии и минеральному питанию и заканчиваем проблемой расселения в кронах, структурой эпифитных сообществ и их экосистемной ролью. На большее мы не претендовали, так как полноценный обзор современной литературы обо всех аспектах биологии эпифитов возможен только в рамках обширной монографии.
Популярные синопсисы