Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Моделирование генетической ассимиляции

Наследование приобретенных признаков в рамках дарвиновского механизма естественного отбора имеет теоретическое обоснование благодаря концепции генетической ассимиляции. В настоящей статье проведено моделирование основных эффектов взаимодействия между обучением (в качестве приобретенного признака) и эволюцией. Эти эффекты таковы: 1) Генетическая ассимиляция приобретаемых в результате индивидуального обучения навыков в течение ряда поколений эволюционного процесса. За счет генетической ассимиляции приобретаемые индивидуально навыки становятся наследуемыми; 2) Эффект экранирования, который приводит к тому, что сильное обучение тормозит эволюционный поиск оптимального генотипа, так как оно увеличивает шансы нахождения хорошего фенотипа независимо от генотипа особи; 3) Эффект нагрузки на обучение, который приводит к уменьшению приспособленности особи таким образом, что это уменьшение тем больше, чем больше было изменение фенотипа в процессе обучения. Наличие нагрузки на обучение может приводить к ускорению эволюционной оптимизации.
При моделировании рассматривалась эволюционирующая популяция модельных особей, каждая особь имеет генотип и фенотип, которые представляют собой цепочки символов большой длины L. Имеются оптимумы для генотипов и фенотипов. Генотипы особей оптимизируются путем эволюции, фенотипы оптимизируются путем обучения.
В начале эволюции генотипы особей случайны. Фенотипы особей в начале поколения определяются их генотипами (в рассмотренной модели начальные фенотипы равны генотипам). В течение поколения фенотипы особей приближаются к оптимумам путем обучения. Генотипы в течение поколения не меняются. В конце поколения происходит отбор особей в следующее поколение в соответствии с их приспособленностями, определяемыми конечными фенотипами особей, полученными в результате обучения. По наследству генотипы особей-родителей передаются (с мутациями) особям-потомкам.
Приведем результат компьютерного моделирования генетической ассимиляции. Считалось, что генотипы SG и фенотипы SP определяют минимизируемый параметр E, равный E (SG) и E(SР), соответственно. Приспособленность особи равна
f = exp[–E (SР)] + Er , (1)
где E(SР) – параметр E, определяемый конечным фенотипом особи SР , а большое слагаемое Er существенно ослабляет силу отбора.
В процессе эволюции и обучения минимизировался параметр E, т.е. максимизировалась приспособленность особей. Рассматривались случаи 1) эволюции совместно с обучением и 2) «чистой» эволюции, т.е. эволюции без обучения. Длина цепочек генотипов и фенотипов L равна 100. Численность популяции равна 100 особей. Типичные значения искомых минимумов параметра E составляют –70. Величина Er равна 1013.
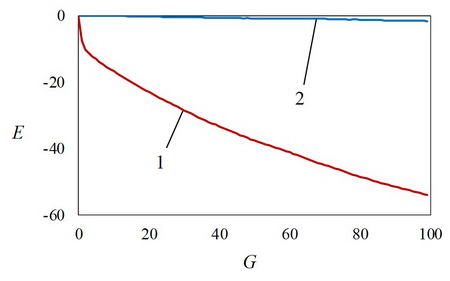
На рис. 1 показана зависимость параметра E, определяемого генотипами особей, E(SG) от номера поколения при наличии обучения (кривая 1) и для «чистой эволюции» в отсутствии обучения (кривая 2). Рис. 1 показывает, что минимизация параметра E при эволюции с обучением значительно эффективнее эволюции без обучения.
Механизм генетической ассимиляции при наличии обучения характеризуется рис. 2. Этот рисунок показывает динамику распределений параметра E для генотипов и фенотипов в первом поколении эволюции при наличии обучения. Видно, что обучение существенно уменьшает величину E и распределение n(E) по фенотипам смещается к меньшим значениям E, а после отбора распределение n(E) по генотипам также смещается, следуя за распределением по фенотипам. Навык уменьшения параметра E «перекодируется» из фенотипов в генотипы, т.е. происходит генетическая ассимиляция.
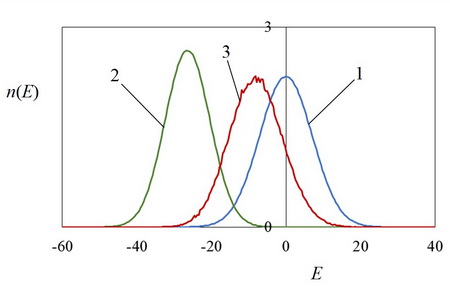
Специфика исследованной генетической ассимиляции связана с большим слагаемым Er в выражении (1). В начальной популяции это слагаемое больше величины exp[–E (SР)], которое может приводить к дифференцированию особей при отборе, так как величина E еще близка к нулю (рис. 2, кривая 1). Поэтому без обучения все приспособленности особей в популяции приближенно равны величине Er , т.е. практически нет дифференцировки особей при отборе. В результате без обучения эволюционный поиск минимумов параметра E сильно ослабевает (рис. 1, кривая 2). Только при наличии обучения величина E (SР) при обучении значительно уменьшается (рис. 2, кривая 2), величины exp[–E (SР)] возрастают, сравниваются с величиной Er , поэтому приспособленности особей становятся различными, и дифференцированный отбор приводит к уменьшению величин E(SGk) в популяции (рис. 1, кривая 1). Итак, генетическая ассимиляция в нашем случае проявляется в том, что обучение направляет популяцию в область облегченного дифференцированного эволюционного отбора особей.
Популярные синопсисы