Последние выпуски
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
- Том 86, № 4. Июль-август 2025
- Том 86, № 3. Май-июнь 2025
Каким будет пополнение?

В популяционной экологии большое внимание уделяется изучению механизмов регуляции популяционного роста. Наиболее эффективным методом исследования таких механизмов и их проявления в динамике численности животных является математическое моделирование. Авторами разработана обобщенная модель формирования пополнения промысловых популяций рыб, дающая полное описание класса зависимостей “запас–пополнение”, основанных на плотностной регуляции динамики популяций. Предлагаемая унифицированная модель обеспечивает возможность количественных сопоставлений (на уровне значений параметров) качественно различных описаний процессов пополнения, основанных на альтернативных гипотезах о механизмах и факторах этих процессов.
Проблема популяционной динамики является одной из ключевых фундаментальных проблем экологической теории. Многообразие и непостоянство факторов и механизмов, контролирующих процессы популяционных изменений, зачастую затрудняет их интерпретацию. Тем не менее все факторы, воздействующие на популяционную динамику живых организмов, можно разделить на экзогенные, т.е. внешние по отношению к популяциям, и эндогенные, т.е. обусловленные структурой и составом самих популяций. К первым относятся климатические и антропогенные воздействия, ко вторым – эффекты плотностной регуляции, проявляющиеся в торможении скорости роста популяции по мере возрастания ее численности. Рост численности вызывает изменение свойств особей, которое выражается в конечном счете в снижении плодовитости, удлинении сроков полового созревания, возрастании смертности и миграционной активности. Динамика многих популяций живых организмов в значительной степени зависит от величины пополнения, т.е. численности вновь нарождающихся генераций. При высокой численности производителей (см. рис. 1) начальная численность молоди также оказывается высокой, что приводит к усилению пищевой конкуренции, повышенной доступности для хищников, паразитов, болезней и пр. В результате смертность молоди возрастает, и итоговая величина пополнения снижается.
Зависимость между численностью родителей и численностью потомков может быть выражена количественно. Такого рода формальные описания широко используются в моделях пополнения, интерпретирующих динамику популяций в контексте саморегуляции. Излишне говорить, что моделей, учитывающих плотностные эффекты в популяционной динамике различных групп растений и животных, к настоящему времени разработано множество. Однако с практической точки зрения особое значение имеют модели пополнения промысловых популяций рыб, поскольку они являются важнейшим инструментом анализа состояния рыбных запасов и управления их промыслом.
Модели пополнения, используемые в настоящее время для описания феномена плотностной регуляции и прогнозирования численности рекрутов, достаточно разнообразны; среди них есть и феноменологичекие (т.е. описывающие изучаемый процесс, не раскрывая механизмы, его контролирующие), и выведенные из некоторых предположений о причинах наблюдаемых популяционных изменений. В рыбохозяйственных исследованиях наиболее употребительными до сих пор остаются модели Рикера и Бивертона–Холта. Модель Рикера (Ricker, 1954) предполагает зависимость смертности молоди рыб от начальной численности генерации и рассматривает в качестве основного фактора, обеспечивающего эту зависимость, хищничество или каннибализм. Модель Бивертона–Холта (Beverton, Holt, 1957) основана на предполагаемой связи смертности молоди с текущей (изменяющейся во времени) численностью генерации, основной причиной которой является конкуренция за пищу.
Авторами статьи разработана модель формирования пополнения промысловых популяций рыб, дающая полное описание класса зависимостей “запас–пополнение”, основанных на максимально общих предположениях о плотностной регуляции динамики популяций. Формулировка модели базируется на решении стохастического дифференциального уравнения динамики генерации на ранних стадиях развития, с параметрами и начальными условиями, зависящими от численности родительского стада. Поскольку модель выведена из первых принципов популяционной динамики (ab initio), в рамках использованного подхода не остается места для феноменологии – все ее свойства, связанные с проявлением эффектов плотностной регуляции, находят должное биологическое обоснование, и наоборот, качественные предположения о биологических процессах проявляют себя в количественных характеристиках модели. В обобщенной формулировке она позволяет прогнозировать реакцию механизмов саморегуляции на все многообразие факторов, контролирующих динамику пополнения, включая условия внешней среды и случайные воздействия. Тем самым появляется возможность разделить эффекты внутрипопуляционной регуляции, предсказанные моделью, и эффекты более сложных механизмов, например, экосистемных, действие которых будет существенно отличаться от предусмотренных моделью сценариев. Одно из главных достоинств предлагаемой унифицированной модели состоит в том, что она позволяет проводить количественные сопоставления (на уровне значений параметров) качественно различных описаний процессов пополнения, основанных на альтернативных гипотезах о механизмах и факторах этих процессов.
Свойства новой модели подробно разобраны на основе анализа ее детерминированной версии, включающей в себя пять управляющих параметров. Один из интересных результатов – взаимовлияние эффектов конкуренции и хищничества/каннибализма при их совместном включении в модельное описание процессов пополнения (рис. 2). Как видно из рисунка, при одновременном действии обоих факторов тип конкуренции влияет и на проявление эффектов хищничества/каннибализма.
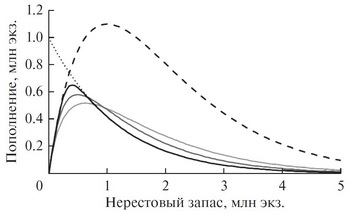
Еще один весьма важный и интересный результат вытекает из анализа предельных форм зависимости “запас–пополнение”, которые позволяют существенно облегчить процедуру оценивания параметров модели. Здесь имеется в виду приближение к так называемой модели “хоккейной клюшки” (hockey stick), в которой кривая пополнения предстает в виде двух связанных линейных сегментов (упрощенный вариант функции пополнения Бивертона–Холта). Результаты исследования предлагаемой обобщенной модели указывают на то, что подобного рода зависимость может определяться нелинейным характером конкуренции и возникает как предельная форма не только функции пополнения Бивертона–Холта, но также и функции пополнения Рикера.
В целом разработанная модель представляет собой достаточно универсальный инструмент, применимый к анализу процессов пополнения как в популяциях рыб, так и в популяциях других живых организмов. Тем не менее, как и любая другая модель, она представляет собой всего лишь упрощенное описание реальной ситуации и оставляет место для дальнейшего развития и детализации.
Популярные синопсисы