Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Гидравлика и никаких нервов: уроки самоорганизации от колониальных гидроидов

Колониальные гидроиды не пользуются центральной регуляцией для синхронизации обменных процессов. Тем не менее успешно осуществляют распределение пищевых частиц между частями организма и доставляют питание к зонам роста. К настоящему моменту механизм перемещения жидкости внутри единой полости колонии описан в общих чертах. В новом исследовании ученые рассчитали объемы жидкости, которые циркулируют в организме модульного животного, определили скорости и протяженность течений, возникающих в стволе колониального гидроида. Расчеты показали, что организация однонаправленного течения, способного доставить пищевые частицы к другим частям колонии, не под силу одному гидранту или любому другому единичному пульсатору. Одномоментное сжатие нескольких пульсаторов, необходимое для создания течения, по-видимому, осуществляется за счет гидравлической синхронизации.
Колониальный гидроид не является унитарной особью, но и не колония различных особей. Это что-то среднее – модульный организм (особь). У колониального гидроида, в отличие от унитарной особи, много одинаковых равноправных частей, и такая особь не сильно страдает от потери значительной части своего тела и легко регенерирует. С другой стороны, в отличие от колоний организмов, весь модульный организм происходит из одного яйца и все части его модульного тела участвуют в общем обмене веществ. У разных модульных организмов наблюдается неодинаковая степень интеграции и регуляции процессов жизнедеятельности. Например, благодаря развитой нервной системе морские перья способны одномоментно реагировать на раздражитель всем своим модульным телом – эти прекрасные животные могут мгновенно спрятаться от назойливых посетителей. А вот у колониальных гидроидов, например у Dynamena pumila, нет единого центра управления процессами обеспечения жизнедеятельности. Таким образом, разные колониальные организмы хорошие модели для изучения роли и соотношения механизмов центральной регуляции и самоорганизации в формировании и функционировании организма.
Н.Н. Марфенин с коллегами с кафедры беспозвоночных МГУ имени М.В. Ломоносова исследуют «физический аспект» существования модульных организмов: Как колониальный гидроид справляется с функцией распределения веществ без централизованного управления? Объектом исследования стал морской колониальный организм (гидроид) Dynamena pumila. Отдельные особи – гидранты – расположены на вертикальных побегах по два напротив друг друга (см. заглавную иллюстрацию и Рис. 1). В свою очередь вертикальные побеги отходят от прямолинейных горизонтальных побегов (столонов) на примерно одинаковых расстояниях. Рост колонии происходит на верхушке столона, от которого ответвляются новые побеги с гидрантами, и на верхушках вертикальных побегов. Как уже говорилось, считается, что у динамены нет единого регуляторного механизма, обеспечивающего синхронизацию процессов жизнедеятельности ее модульного организма. Тем не менее всю колонию объединяет общая (гастроваскулярная) полость, по которой циркулирует жидкость (гидроплазма) с пищевыми частицами, захваченными отдельными гидрантами. Ток гидроплазмы создается пульсациями отдельных гидрантов и поперечным сокращением стенок ствола колонии (пульсаторами). Течение жидкости не имеет постоянного однонаправленного характера (как, например, у гидроида Tubularia) и может быть разнонаправленным как в разных участках колонии одновременно, так и в одном и том же участке в разное время. Целью данной работы стало вычисление объемов гидроплазмы, перемещаемых по колонии гидроида, на основе данных видеорегистрации и математического моделирования.
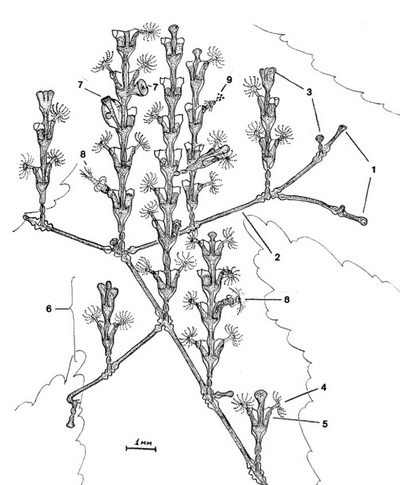
Колонии гидроидов собрали в Кандалакшском заливе Белого моря. В лаборатории на искусственном субстрате из фрагментов собранных колоний, составлявших в новых организмах материнские побеги, вырастили экспериментальные организмы. Было исследовано семь прямолинейных неразветвленных колоний, в которых по числу гидрантов доминировали материнские побеги, тогда как остальные побеги включали всего по несколько пар гидрантов каждый. Для экспериментов составили схемы выращенных колоний. Под микроскопом визуальным методом определяли направление, насыщенность пищевыми частицами и протяженность неразрывного гидроплазматического течения. Пульсации различных участков колонии регистрировали с помощью видеосъемки. Для определения объемов жидкости разработали и применили упрощенную схему внутренних полостей побега (например, столон рассматривали как цилиндр, полость желудка гидранта в виде двух усеченных параболоидов и т.д.) и замеряли диаметры просвета полостей в разных «режимах» работы пульсаторов.
Основными результатами работы стали:
некоторые морфометрические характеристики исследованных колоний;
регистрация протяженности течений и их скоростей;
вычисленные объемы жидкости, которые продуцирует каждый отдельный участок-пульсатор, т.е. разница в объемах жидкости в состоянии сжатия и расширения каждого пульсатора, так называемый «рабочий объем»;
расчет объемов гидроплазмы однонаправленного течения в ближайшем к побегу участке столона (модуле столона);
Представленная авторами временная схема динамики перемещения жидкости показывает, что большую часть времени в теле колониального организма нет поступательного движения жидкости. Однако периодически возникает сильное однонаправленное течение, которое начинается от материнского (старого) побега к верхушке столона (зона роста колонии). Скорость течения набольшая в участке столона, находящегося у материнского побега, и наименьшая у верхушки столона.
Рабочие объемы довольно различны в разных модулях столона. При приблизительно одинаковых размерах просвета полости столона (в покое) наибольшая амплитуда сжатия, т.е. рабочий объем, оказалась у верхушечного модуля столона, несколько меньше у второго модуля, а начиная с третьего модуля амплитуды приблизительно одинаковы у всех последующих участков столона. В побеге тоже наблюдается разница объемов связанная с положением относительно верхушки роста побега: объем полости вблизи верхушки роста всегда больше, чем на участке побега, находящегося ближе к столону. В статье приведены объемы всех участков побега. Например, вычисленный максимальный рабочий объем модуля побега (два гидранта с ножками+междоузлие) составляет около 6 790 197 мкм3.
Исследователи рассчитали, что для создания локального течения на участке столона между побегами необходима жидкость в объеме от 1 542 315 мкм3 (слабое течение) до 50 147 268 мкм3 (сильное течение). Получается, что сильное течение может создать только одновременное сжатие нескольких пульсаторов. При этом оказалось, что визуально регистрируемое течение протяженностью 3-4 модуля столона, на самом деле (согласно математическим расчетам по вычисленным объемам) составляет не больше двух модулей. Из этого следует, что течение создается не одним только материнским побегом, но и другими побегами. Можно сравнить возникновение однонаправленного течения с передачей эстафеты жидкости по столону от материнского побега следующим побегам, а от них к следующим и так до верхушки столона. Хотя ранее другими исследователями была описана такая картина, оставалось неясно сколько же пульсаторов могут начать и поддерживать «эстафету». Настоящее исследование показало, что для рождения сильного однонаправленного течения необходимо одномоментное сжатие нескольких пульсаторов.
Как же достигается синхронизация независимых пульсаторов? Авторы предполагают, что их исследование косвенно свидетельствует о наличии гидравлического механизма синхронизации. Дело в том, что гидранты (надо полагать, что и другие пульсаторы колонии) сокращаются независимо друг от друга, но собственный ритм зависит от расширения полости пульсатора в результате наполнения жидкостью. Так, по-видимому, возникает авторегуляторный цикл – поступающая в побег в достаточном объеме жидкость от соседнего побега расширяет стенки его полостей и вызывает одновременное сжатие нескольких пульсаторов. На мой взгляд (КП), можно было бы привести статистику пульсаций (сокращение, расслабление и расширение) различных пульсаторов модульного организма и наложить ее на временную схему возникающих течений и тем самым прямо показать связь между течениями и синхронизацией сокращений.
Популярные синопсисы