Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Расселение тихоокеанских моллюсков как результат движения материков

Известный биогеограф А. И. Кафанов из Института биологии моря ДВО РАН представил краткий итог многолетней работы по изучению расселения двустворчатых моллюсков Тихого океана от начала кайнозоя (65 млн лет назад) и до современности. В прошлых номерах журнала А. И. Кафанов высказал свое мнение по поводу механизмов формирования крупных биогеографических подразделений: биогеографические регионы ранга области и подобласти складываются под влиянием глобальных изменений климата, дрейфа материков и других планетарных явлений (см.: О предмете биогеографии). В своей новой статье Кафанов показывает, как в течение кайнозоя под влиянием планетарных явлений сформировалась Северотихоокеанская биогеографическая область. Ясно, что подобному биогеографическому анализу предшествовала долгая работа по сбору данных. Из списка литературы и введения мы можем узнать, что составление таксономических списков двустворок, переопределение видов и приведение их в соответствие с кодексом зоологической номенклатуры заняло у автора около 10 лет. Первые работы со списками видов начали публиковаться в 1996 году, и теперь автор имеет в своем распоряжении отредактированные списки для Японии, Дальнего Востока и североамериканского побережья Аляски и Калифорнии (например, для Дальнего Востока список включает 793 валидных вида, относящихся к 225 родам).
Далее предстояло решить, какие временные (хронологические) единицы использовать в данной работе. Действительно, если рассматриваешь историю, то нужно по одной из осей откладывать какое-то время. Обычно в исторической биогеографии пользуются веками (ярусами) и зонами (хронами) (см.: Хронология далекого прошлого). Однако для Дальнего Востока эти временные единицы не подходили. Еще раньше в специальной статье в Палеонтологическом журнале (2003 г., № 6) Кафанов обращал внимание на плачевное состояние стратиграфии этого района. К моменту опубликования статьи в Палеонтологическом журнале строгого определения ярусов и зон кайнозоя на Дальнем Востоке не было. Из-за этого пришлось воспользоваться не геохронологическими подразделениями ранга зон, а геологическими единицами типа свит и оперировать списками видов, найденных в каждой свите. Для кайнозойских отложений побережья Тихого океана известно около 500 таких свит с фауной двустворчатых моллюсков, именно они и стали основой биогеографического анализа. По сходству фаун отдельные свиты разделены на видовые комплексы. Всего автор различает 11 комплексов моллюсков, последовательно сменявших друг друга во времени. Эта огромная, но вспомогательная для данной задачи работа была проделана для того, чтобы понять, какие комплексы каким временным единицам (зонам, ярусам) соответствуют.
А для биогеографического исследования все же использовались не комлексы, а свиты. Причем были выбраны такие свиты, в которых двустворок не меньше десяти видов. Таких свит примерно четверть из всех известных для кайнозоя Тихого океана. Затем был проведен кластерный анализ выбранных списков видов, то есть списки сгруппировали по сходству. Какие же в результате получились группы?
Я схематично изобразила развитие биогеографических подразделений Северной Пацифики в течение кайнозоя так, как это описано в статье (гигантские кладограммы свит можно посмотреть в оригинальной статье).
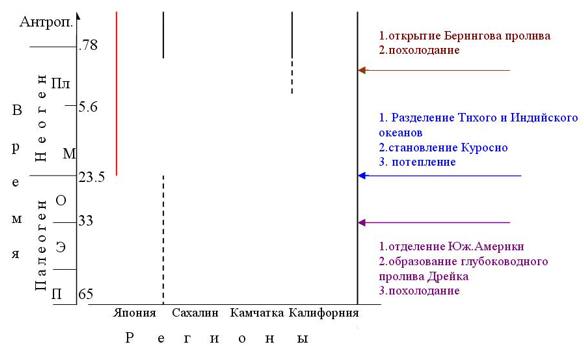
На этой схеме показано, что рассматриваются отложения четырех регионов: Японии, Сахалина, Камчатки и Калифорнии. По вертикальной оси отложено время и буквами отмечены яруса палеогена и неогена: палеоцен, эоцен, олигоцен, миоцен, плиоцен, и выше антропоген. Вертикальные линии обозначают границы биогеографических подразделений. Красной линией показано обособление Северотихоокеанской области, сплошной черной линией обозначены границы подобластей, а пунктиром – провинций. Итак, в раннем палеогене Северотихоокеанская область в ее современных очертаниях еще не обособилась. Видно, что Японские двустворки представляли собой обособленную фауну. Больше всего эта фауна похожа на тропических моллюсков древнего океана Тетис, а не на своих Камчатских соседей.
Произошедшее на рубеже эоцена и олигоцена отделение Южной Америки от Антарктиды, сопровождавшееся образование глубоководного пролива Дрейка и похолоданием северных побережий Тихого океана, не повлекло заметных изменений в биогеографической системе. Зато позже из-за разъединения Тихого и Индийского океанов японские виды двустворок стали больше соответствовать другим Тихоокеанским видам, таким образом Северотихоокеанская область обособилась.
Потепление и ровный климат в среднем миоцене способствовали относительно равномерному распределению видов по этим акваториям: провинций и подобластей еще не было. Постепенное обособление азиатской и американской фаун началось в только в позднем миоцене. В этом веке еще существовали активные миграции (точнее — прохорезы) между Камчаткими морскими бассейнами и Американскими. Только позже, в среднем плиоцене, фауна американских двустворок обособляется, формируя Орегонскую или Орегоно-Ситканскую подобласть. Чуть позже в связи с открытием Берингова пролива разделяются Берингийская и Японо-Маньчжурская подобласти.
Так, по мнению Кафанова, постепенно сформировалась Северотихоокеанская область с тремя подбластями. Таким образом Кафанов относительно успешно совмещает глобальные климатические и географические события с появлением и дифференцировкой одной из биогеографических областей современности.
Как указал сам автор статьи, здесь впервые кратко и на русском языке публикуются результаты многолетних исследований, опубликованных прежде в малодоступных журналах и на английском. Вместе с тем у читателей этой русской публикации наверняка возникнет немало вопросов. Например, почему образование глубоководного пролива между Антарктидой и Южной Америкой мало повлияло на фауны олигоценовых моллюсков. Возможно, дело тут в недостатке материала по олигоценовым отложениям, а возможно и в чем-то другом — русским читателям можно только гадать.
Кроме того, почему обособление американской фауны от Камчатской началось раньше, чем Японской от Камчатской? Какие из глобальных событий — а названо их много — были ведущими в формировании области и подобластей? Читателю наверняка будет интересен более подробный разбор биогеографических механизмов.
См. также:
А. И. Кафанов. Континуальность и дискретность живого покрова: проблема масштаба // Журнал общей биологии. 2006. Том 67. № 4. Стр. 311-313.
А. И. Кафанов, В. А. Кудряшов. Классики общей биогеографии: биобиблиографический указатель.
К. Ю. Еськов. Историческая биогеография.
Популярные синопсисы