Последние выпуски
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
- Том 86, № 4. Июль-август 2025
- Том 86, № 3. Май-июнь 2025
Пестики и тычинки: запрещенные темы
Эволюционные пути, приводящие к формированию цветка с одним просто устроенным пестиком, могут быть разные. Либо это сокращение исходно многочисленных пестиков до одного, либо упрощение изначально сложно устроенного пестика. Согласно своему происхождению внешне сходные просто устроенные пестики описываются с помощью разных наукообразных терминов – как мономерный гинецей и псевдомономерный гинецей («псевдо», поскольку гинецей проходит через стадию срастания пестиков друг с другом). Для разрешения проблемы различения между собой этих типов строения женской сферы цветка (=гинецея) задействованы классические и современные методы исследования морфологии структур: сравнительная анатомия, изучение морфогенеза, молекулярно-генетический подход. Перспективны также исследования эволюционных запретов некоторых преобразований. Однако однозначного решения пока нет. Сложности носят не только биологический характер, но и отчасти терминологический. Жестко привязывая термины к явлениям (структурам), исследователи ограничивают развитие понимания биологического явления (структуры) как такового.
Строение гинецея имеет для эволюционной ботаники огромное значение. Долгое время (до появления молекулярной филогенетики) именно строение репродуктивной части организма было основой для установления эволюционных процессов и родственных отношений растений. Из всего многообразия морфологии цветков ученые пытаются выстроить последовательные в эволюционном смысле ряды преобразований. Для этих целей необходимо иметь разработанную терминологию и понятийный аппарат. Однако даже при детальном всестороннем анализе морфологических структур имеются случаи, когда разграничение между собой отдельных (часто терминальных) стадий независимых эволюционных линий на практике крайне сложно. В статье заведующего кафедрой высших растений МГУ им. М.В. Ломоносова Д.Д. Соколова обсуждается один из таких сложных моментов эволюционной морфологии цветковых растений.

Немного вводной части. Умным словом плодолистик называют элементарную единицу женской сферы в цветке. Есть веские основания думать, что плодолистики всех растений гомологичны между собой, и менее веские (хотя значительные) – что они произошли от какой-то листовидной структуры, что и отражено в русском термине «плодоЛИСТИК». На самом деле, никто не знает, как возник плодолистик (см. подробнее популярный синопсис к статье Д.Д. Соколова и А.К. Тимонина Морфологические и молекулярно-генетические данные о происхождении цветка: на пути к синтезу). Если плодолистики свободные, то каждый из них = пестик. Если они срослись между собой, то все это вместе = 1 пестик. Поэтому, когда в определителе растений читаем, что у растения 1 пестик, это намеренное уклонение от обращения к вопросу о гомологиях. Совокупность плодолистиков называется гинецей. Если плодолистиков больше одного, то гинецей полимерный (в противном случае – мономерный). Различают несколько типов полимерного гинецея (основные из них - апокарпный и ценокарпный, рис. 1). В зависимости от того, как устроена и расположена завязь относительно других частей цветка, ее именуют верхняя, полунижняя или нижняя (см. рис. 1).
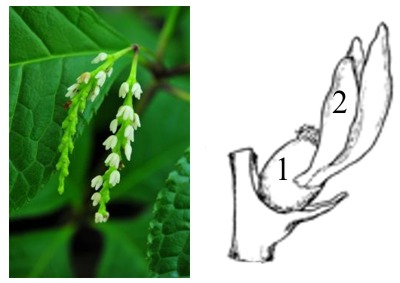
Проблема, обсуждаемая в данной статье, вызвана затруднением различить между собой стадию цветка с одним плодолистиком (см. рисунок заглавной иллюстрации) двух принципиально разных эволюционных рядов. Такой цветок мог получиться в ходе эволюции сокращением числа плодолистиков у изначально апокарпного и полимерного (несколько плодолистиков) цветка, т.е. у которого плодолистики были свободными. Этим путем получается мономерный гинецей. Другим путем может быть упрощение изначально полимерного, но ценокрапного гинецея, т.е. у которого плодолистики еще до упрощения были сросшимися. Так получается псевдомономерный гинецей. Гордиев узел попробовал разрубить Г.Б. Кедров. На основании того, что у всего морфологического разнообразия цветков покрытосеменных нет случаев апокарпного полимерного гинецея с нижней завязью, он предположил наличие запрета на это сочетание признаков. Следуя этой логике, если в цветке один плодолистик и нижняя завязь – то это – обман зрения. На самом деле, плодолистиков несколько, но все, кроме одного, сильно редуцированы и приросли к нормально развитому (псевдомономерия). Для появления гипотезы вполне достаточно анализа морфологического разнообразия, но для ее развития и проверки необходимо найти механизм «запрета». А вот это очень проблематично, хотя на пути поиска подобных механизмов ожидается много открытий.
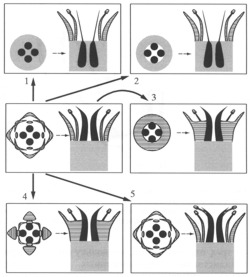
Дмитрий Соколов подробно описал проблематичные ситуации и рассмотрел возможные причины отсутствия цветков со свободными многочисленными плодолистиками и нижней завязью. Для этого были представлены и критически рассмотрены возможные теоретические пути становления нижней завязи на базе гинецея со свободными плодолистиками (рис. 2):
1) погружение завязи каждого плодолистика в отдельности в особое углубление цветоложа;
2) погружение гинецея в целом в единое углубление цветоложа;
3) прирастание к гинецею трубки, образованной сросшимися основаниями прочих элементов околоцветника и/или тычинок);
4) прирастание к гинецею или отдельным плодолистикам элементов околоцветника и/или тычинок в индивидуальном порядке без их предварительного срастания между собой;
5) появление на отдельных плодолистиках или ценокарпном гинецее тычинок или элементов околоцветника de novo (чисто умозрительный вариант).
Автор исследования приходит к заключению, что отсутствию апокарпных цветков с нижней завязью могут способствовать несколько причин биологического характера, но и, отчасти, особенности того, как ботаники определяют понятие «ценокарпия». Например, при первом или втором пути преобразований, завязи, которые опускаются и развиваются в пространстве цветочной трубки (сросшиеся основания чашелистиков) до эволюционного момента их прирастания к тканям цветочной трубки называются верхними. Однако после этого момента, гинецей автоматически будет именоваться (но является ли?) ценокарпным. Детальный анализ хода развития цветков обнаруживает также и препятствия морфогенетического характера. Например, невозможно радиальное срастание отдельных плодолистиков с тычинками (это необходимо для того, чтобы завязь оказалась в нижнем положении) без срастания их между собой (ведь плодолистики должны оставаться свободными!).
В заключительной части статьи подводятся итоги проделанной работе. Особое внимание уделяется необходимости новых подходов для решения спорных вопросов эволюционной морфологии. Среди них и перспективы исследований морфогенеза цветков и пересмотр отношения к понятийному аппарату, который, конечно, дает основу для начала движения в науке, но без переосмысления и обновления тормозит рождение и развитие идей.
Автор синопсиса признательна Дмитрию Дмитриевичу Соколову за коррекцию и терминологическую правку текста синопсиса.
Подробнее по теме читайте научно-популярную статью Д.Д. Соколова ''Эволюционная ботаника: так много вопросов на поставленные ответы.''
Популярные синопсисы