Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Ранняя история Metazoa – взгляд палеонтолога
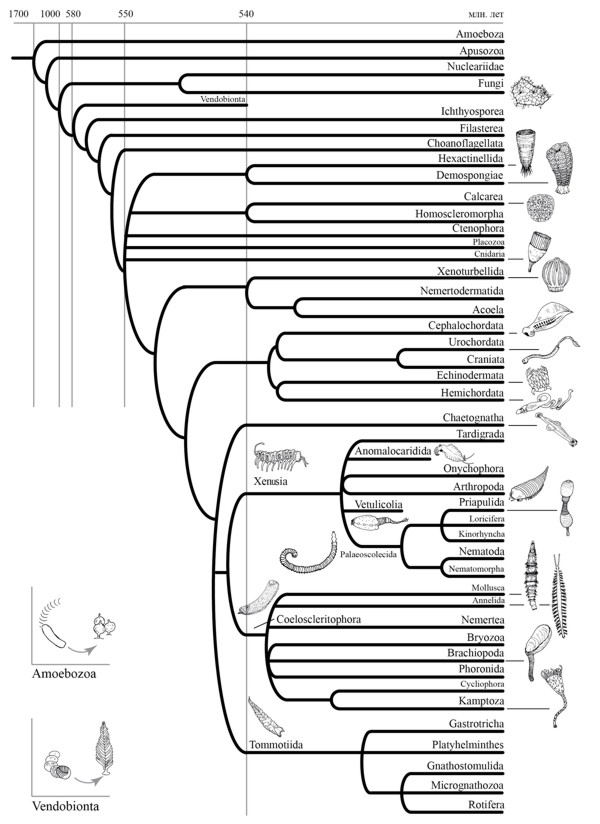
В статье палеонтолога Андрея Журавлева из МГУ имени М.В. Ломоносова рассмотрены современные представления о филогении многоклеточных животных (Metazoa). За последние 10-15 лет система родственных отношений Metazoa претерпела кардинальные изменения. Стремительно развивающиеся молекулярные методы, потрясающие и интенсивные палеонтологические исследования ранней истории многоклеточных открывают для нас мир, отсутствие которого так смущало Ч. Дарвина и его единомышленников.
Успехи молекулярной биологии, заставившие переосмыслить базовые представления о родстве и путях эволюции крупных групп («типов») многоклеточных животных, с большим вниманием были встречены палеонтологами, чем неонтологами (т.е. биологами, изучающими современный органический мир). Это неудивительно: именно ископаемая летопись является надежным проверочным тестом для молекулярно-биологических гипотез. Палеонтологическая летопись показывает, что между различными «типами», объединяемыми ныне в ветвь Ecdysozoa (линяющие животные), – членистоногими, онихофорами, тихоходками, приапулидами, волосатиками – существовало множество переходных форм, обитавших в морской среде и вымерших еще в раннем палеозое (см. заглавную иллюстрацию).
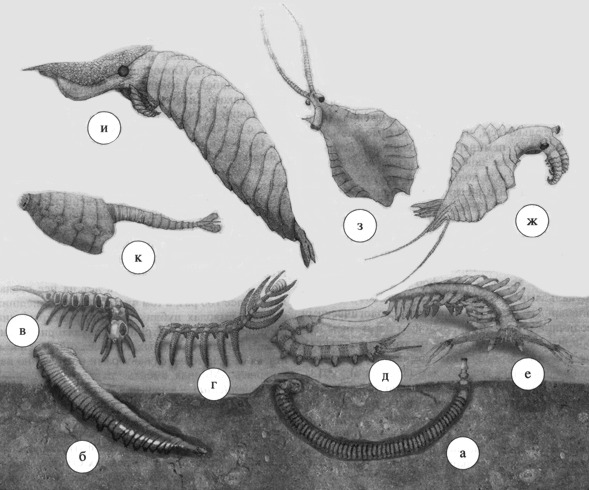
В первую очередь это были ксенузии (Xenusia) – весьма мозаичная группа, включающая червеобразных животных с лобоподными, телескопически втягивающимися ходными конечностями (как у современных тихоходок и онихофор), терминальными ротовым и анальным отверстиями и трёхслойной кутикулой, на нижней поверхности которой сохранились закономерным образом расположенные полости, где при жизни располагались коллагеновые волокна (как в кутикуле круглых червей). Ротовое отверстие располагалось на кончике хоботка, который у некоторых форм был втяжным (как у головохоботных червей). Облик этих организмов вполне соответствует гипотетическим исходным формам, которые должны были существовать, судя по онтогенезу (индивидуальному развитию) онихофор. Переход части ксенузий к роющему образу жизни дал начало головохоботным червям, у древнейшей группы которых – палеосколецид – сохранилась кутикула, состоявшая из точно таких же склеритов (фосфатизированных пластинок), как у ксенузий, и редуцированными в различной степени лобоподными конечностями, крайним случаем редукции которых были сенсорно-железистые придатки – тубули, расположенные двумя отчётливыми вентро-латеральными (т.е. по бокам брюшной стороны) рядами. Другие группы ксенузий дали онихофор, перешедших к наземному существованию, тихоходок – мейобентосную группу и аномалокаридид – крупные нектонные формы, утратившие ходные конечности и вымершие в палеозое (рис. 1). Остатки каких-либо животных, которые даже с очень большой натяжкой можно было бы рассматривать как промежуточные формы между членистоногими и кольчецами, в ископаемой летописи полностью отсутствуют.
Древнейшие Deuterostomia – единственной ветви Bilateria (двусторонне-симметричные животные), не оспариваемой ни одной из дисциплин, – хотя и в меньших подробностях, позволяют воссоздать их раннюю историю и указывают на наличие подвижных двусторонне-симметричных форм в начале становления как хордовых, так и гемихордовых и иглокожих.
Сложнее понять раннюю эволюцию Lophotrochozoa, поскольку в отличие от других ветвей Bilateria их древнейшие остатки представлены исключительно скелетными ископаемыми. Однако именно общность скелетных микроструктур у моллюсков, брахиопод и мшанок, не встречающихся у других Metazoa, указывает на то, что среди древнейших Lophotrochozoa были близкородственные формы, часть которых являлась сидячими сестонофагами (т.е. фильтраторами - ханцеллорииды, томмотииды), часть – подвижными эпибентосными детритофагами (халькиерииды, махайридии). Все эти формы имели многостворчатые известковые или фосфатные склеритомы (твердые компоненты скелета), редуцировавшиеся в различной степени за время эволюции.
Среди более древних – докембрийских групп выделяются ископаемые следы группы Gaojiashania – Myxomitodes – Horodyskia, оставленные на морском дне 1,7–0,55 млрд лет назад, и гигантские вендобионты, существовавшие 0,58–0,43 млн лет назад. Первые из них напоминают следы, оставляемые временными агрегационными колониями Amoebozoa (прежние акразиевые слизевики), и цепочки плодовых тел тех же организмов. Вендобионты – представляют собой крупные (до одного-двух метров длиной/высотой) организмы с соотношением площади тела и его объёму на уровне 1•100–1•102 мм-1, то есть как у осмотрофных мегабактерий и ниже, тело которых было пронизано системой бифуркирующих и уменьшающихся к периферии в диаметре каналов (рис. 2). Вендобионты могли быть приспособлены к осмотрофному питанию в условиях океана, насыщенного растворённым, дисперсным и коллоидным органическим веществом, каким и был океан эдиакарского периода. Достоверных остатков каких-либо групп организмов, которых можно было бы отнести к многоклеточным животным, в докембрийских отложения не обнаружено.

Однако данные молекулярной биологии показывают, что комплекс ключевых генов Metazoa (многоклеточных), задействованных в эмбриональном развитии, половой репродукции и апоптозе (саморазрушении клетки), а именно, отвечающих за факторы регуляции транскрипции, склеивание и поляризацию клеток в тканях, коммуникативные межклеточные системы, клеточную дифференциацию, а также гены, производящие нейромедиаторы (вещества-посредники для передачи сигнала от нервной клетки к другим клеткам), есть у воротничковых жгутиконосцев и других одноклеточных Opisthokonta (Ichthyosporea, Filasterea, Chytridiomycota, Apusozoa), причём у грибов и воротничковых жгутиконосцев часть этого генного комплекса (от 30 до 50%) могла быть вторично утрачена. Этот генный комплекс включается на стадии образования колоний и служит, например, для синхронизации деления клеток. Вендобионты могли быть одной из ранних групп многоклеточных Opisthokonta, приспособленных к определённым условиям и не оставивших потомков. В свою очередь предковые Amoebozoa могли быть группой исходной для Unikonta (см. заглавную иллюстрацию).
В совокупности современные данные молекулярной биологии, палеонтологии и сравнительной эмбриологии/анатомии, вышедшие на новый уровень с внедрением новых методов микроскопии, позволяют из всех предположений о происхождении Metazoa наименее вероятными считать гипотезы организма, подобного гастрее, или иного планктонного предка. Более вероятно, что общий предок Bilateria был подвижным эпибентосным животным, а стремительная диверсификация Metazoa, охватившая конец эдиакарского – начало кембрийского периодов (около 40 млн лет) была реальным событием, которому предшествовала длившаяся более миллиарда лет сборка метазойного генома у общих предков Opisthokonta и Unikonta в целом.
Полный текст статьи можно найти здесь.Популярные синопсисы