Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Склонность к «незаконному браку»* передается по наследству
Классическое понимание формирования и передачи наследственной информации предусматривает внутривидовую репродуктивную целостность. Исследования роли горизонтального переноса генов (между неродственными формами) значительно трансформировали такую концепцию. В последнее время активно изучаются примеры межвидовых взаимодействий на генном уровне, и формулируется новая концепция надвидовых генных систем. Согласно последней, степень взаимовлияния организмов обуславливает тот или иной уровень объединения их генетических систем. Различают три типа надвидовых систем наследственности: метагеном (совокупность наследственных факторов микробного сообщества), симбиогеном (функционально интегрированная система генов, которые принадлежат тесно взаимодействующим партнерам симбиоза) и хологеном (система наследственности симбиогенно возникшего организма). С процессами интеграции наследственных систем симбиозов наблюдается эволюция передачи симбионтов в поколениях хозяина: от псевдо-вертикальной через внешнюю среду к наиболее надежной – через половые клетки. Возникновение, способы формирования, механизмы эволюции надвидовых генетических систем изучаются на примерах экологически, функционально взаимосвязанных организмов.
В настоящее время разделение на наследственные и средовые факторы при формировании фенотипа организма представляется слишком размытым и требующим многих уточнений. Стало очевидным, что фенотип эукариот определяется сложной системой взаимодействий генетических, эпигенетических и средовых факторов. Более того, у высших организмов, например, помимо собственного наследственного материала, важную роль в формировании фенотипа играет совокупный геном симбиотического микробного сообщества (см., например, статью на элементах). Поэтому важно сформулировать концепции надвидовых систем наследственности. Систематизация представлений о надвидовых генетических системах представлена в обзоре Н. А. Проворова и И. А. Тихоновича из Санкт-Петербурга.

Разные эволюционные фазы интеграции геномов и генных сетей можно проследить на примерах геномов симбиотических бактерий. Например, кишечная палочка Escherichia coli не имеет специфических генов, ответственных за симбиотические взаимодействия. Однако у ее ближайших родственников, патогенных шигелл, имеются специальные плазмиды (небольшие свободные кольцевые ДНК) необходимые для обеспечения симбиоза. Интересно, что у зависимых от симбиоза микроорганизмов генетическая информация, ответственная за симбиотические связи, находится в дополнительной внехромосомной части генома. Причем эта часть генома у шигелл-симбионтов значительно увеличена по сравнению со свободноживущими родственными формами. Таким образом, эволюция надвидовых систем наследственности проходит на основе многокомпонентного генома, когда наследственная информация может располагаться в различных структурных элементах генома. Дальнейшая эволюция генома связана с утратой микросимбионтами, обитающими в стабильной среде, части генов, ответственных за некоторые метаболические пути. Не смотря на далеко зашедшую интеграцию, изменения геномов симбионтов вплоть до этого этапа носят функциональный характер, т.е не происходит структурной перестройки как таковой (см. ниже). Горизонтальный перенос генов и реорганизация собственного генома микросимбионтов являются основными механизмами, обеспечивающими эволюцию на этих этапах формирования общих систем наследственности.
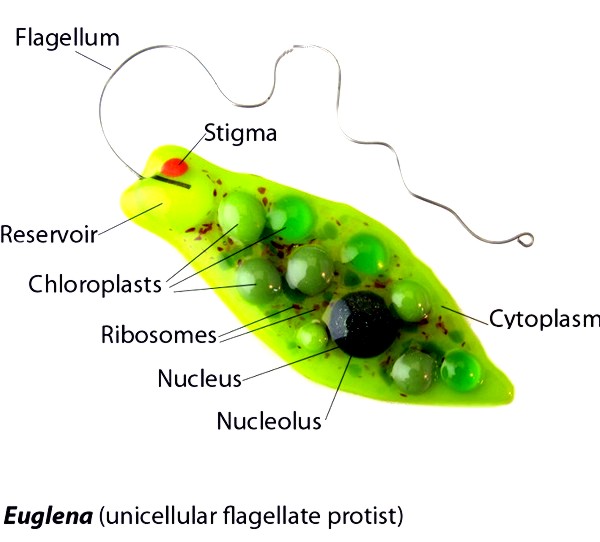
На примере внутриклеточных органелл можно видеть, что эволюция в направлении большей интеграции приводит к структурному объединению геномов, когда часть собственных генов органелл переносится в ядерную ДНК хозяина. Механизм переноса представляется следующим: дупликация генов – фиксация новой копии гена эндосимбионта в геноме хозяина – утрата исходной копии. В дальнейшем перенесенные гены в геноме хозяина эволюционируют относительно независимо от собственного генома эндосимбионта и могут приобретать новые функции. Экстремальным результатом перестройки в направлении переноса генов эндосимбионтов в ядерную ДНК хозяина может быть даже редукция органелл. Так, например, у некоторых эвглен и альвеолят неоднократно происходиоа утрата пластид, однако в их ядерных геномах сохранились многочисленные копии бактериальных генов, которые приобрели новые функции, не связанные с поддержанием этих органелл.
Исследования надвидовых систем наследственности показывают механизмы коэволюции неродственных организмов на самом глубинном – генном уровне. Авторы обзора видят в исследованиях таких систем ключ к пониманию связи процессов макро- и микро-эволюции.
«незаконный брак»* - одно из значений греческого слова «симбиоз».
Читайте также:
обзор по эволюции симбиотических отношений в ‘‘Журнале общей биологии’’ и
статью на Элементах ‘‘Метаболическая интеграция организмов в системах симбиоза’’
Популярные синопсисы