Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Трудный путь от камуфляжа к боевой раскраске: математический подход

Среди животных, имеющих надежные средства защиты (сильнодействующие яды, горький вкус, зловонные вещества, острые шипы), принято предупреждать хищников о своей несъедобности. Выгодно это обеим сторонам: первые не желали бы быть ошибочно съеденными (или хотя бы поврежденными), а в планы вторых не входит наглотаться всякой гадости. Вполне естественно, что в процессе эволюции ядовитые и несъедобные животные приобрели предупреждающую окраску — яркую, часто пёструю, — демонстрирующую, что трогать их нельзя (рис. 1). Однако не вполне очевидным представляется момент зарождения и распространения предупреждающей окраски, потому что самые первые, еще малочисленные, броские особи подвергаются максимальному риску — хищник уже хорошо видит этих животных, но еще не осведомлен об их несъедобности. А посему может случайно их истребить, так и не дав распространиться в популяции жертв спасительному яркому качеству. Иными словами, на ранних этапах эволюционного перехода от защитной (криптической) окраски к броской «предупреждающей» (апосематической) животные должны сталкиваться с проблемой временного снижения приспособленности — преодолевать порог отрицательного отбора. Имитационная математическая модель, учитывающая такие параметры, как цена несъедобности, цена предупреждающей окраски, цена комбинированной окраски, цена атаки хищника, вероятности обнаружения хищником жертв с различной окраской, скорость обучения и длительность памяти хищника — позволяет представить развитие эволюционных процессов становления предупреждающей окраски.
Многие живые организмы имеют защитную (криптическую, от греч. krypte — тайный, скрытый) окраску, соответствующую по цветовой гамме окружающему ландшафту или его элементам (рис. 1). Скучный монохромно-криптический мир, впрочем, достаточно задорно представлен в старой детской песенке — «Оранжевое море, оранжевое небо, оранжевая зелень, оранжевый верблюд...» Оранжевые мамы охраняют (увы, не цветные) сны оранжевых младенцев, отгоняя назойливых оранжевых мух и оранжевых же ос. Им помогают оранжевые птицы, которые охотятся на насекомых.
Очень часто птицы испытывают глубокое разочарование, склевав вместо вкусной мухи ядовитую осу или противного зловонного клопа. Последним это также крайне неприятно. Налицо совпадение интересов «верхов» и «низов» трофической цепи.
Назрела эволюционная ситуация. Пора переходить к предостерегающей (апосематической, от греч. ароsematio — указываю, предвещаю) окраске. В процессе эволюции осы опоясывают тело контрастными ленточками, а иные клопы, так и вообще, рядятся в красно-чёрный мундир «солдатика» (рис. 1). Птицы же вынужденно обучаются не есть все броское и яркое.
Нелегко приходится обычным мухам-жертвам, но и они коэволюционируют под прессингом безжалостного птичьего клюва: выживают наиболее юркие и плодовитые.
Впрочем, направление эволюционного развития не следует олицетворять с единым «перстом указующим» — это, скорее, растопыренная ладонь фантастического «стопальцевика», причём очень многие большие пальцы загибаются назад к запястью — в небытие…
Порою воспользоваться уже имеющимся в данной местности трафаретом, сигнализирующем об опасности, проще и дешевле, чем изобретать нечто оригинальное. И вот некоторые «ленивые» мухи пошли по другому пути, остроумно раскрасив тело по образцу ос и пчёл (мюллеровская мимикрия, мимикрия Бейтса). При этом, по мнению некоторых испанских ученых, сходство рисунка необязательно, важнее яркость и цвет — желтый, красный, синий, чёрный и белый. Хищники хорошо запоминают блок расцветок опасных жертв, причем, чем шире круг последних, тем быстрее потенциальные агрессоры обучатся не трогать никого из них (см. также: мимикрия на «Элементах»).
В общем, несъедобным животным выгодно быть яркими, чтобы предупреждать хишников о своей несъедобности. А хищникам выгодно не есть ярких, чтобы не отравиться. На этом фоне могут появляться «хитрые» животные, которые, хоть и съедобные, но все равно яркие, потому что хищники и их есть не станут. Вопрос лишь один — как такая адаптация, как апосематическая окраска, могла появиться в ходе эволюции. По мнению многих учёных, имеет место «эволюционный парадокс» — на раннем этапе эволюции любая особь, отличающаяся от камуфляжной, моментально поедается хищником, то есть существует область непреодолимого отрицательного отбора. Иногда образно говорят о «пересечении оврага в адаптивном ландшафте».
Автор обсуждаемой статьи В.И. Грабовский (Центр по проблемам экологии и продуктивности лесов) предлагает математическую модель, объясняющую, как этот барьер может быть преодолен. В модели имитируется случайный дрейф генов (мутаций) в сторону цветовых отклонений от стандартной формы. Как и всякая модель, эта модель основывается на ряде дополнительных допущений.
В модели приняты следующие исходные условия:
- в популяции жертв 1000 однополых особей, размножаются бесполым путём, для простоты;
- популяция изначально полностью криптична (все особи одинаково «оранжевы»).
Время в модели исчисляется в непересекающихся поколениях жертв, или тактах. Каждая особь на каждом такте времени может мутировать с вероятностью 0,01 мутаций на особь за поколение. При мутации изменяется один из двух параметров: съедобность или окраска. Причем по обоим направлениям возможны как прямые, так и обратные мутации. Т.е. исходно съедобные жертвы могут стать несъедобными, а потом, наоборот, потерять приобретенные свойства несъедобности и снова стать съедобными.

Окраска в модели представлена в трех вариантах: камуфляжной (исходный вариант), предостерегающей (измененной), или комбинированной, когда особи демонстрируют предостерегающую окраску лишь будучи обнаруженными хищником. Окраска определяется двумя генами. Камуфляжный, или исходный, немутантный, фенотип, определяется состоянием «0-0», предостерегающая окраска имеет «генотип» «1-1» (две мутации), а комбинированная ─ «0-1» или «1-0» (только одну мутацию).
В природе некоторым видам несъедобность дается даром, в виде химических соединений кормовых растений — безобидных для жертвы, но ядовитых для позвоночных хищников, но в данной модели ядовитость предполагается платной (Cpois) — мутацией, снижающей плодовитость. Живтоное платит за несъедобность снижением плодовитости. Это одно из допущений модели. Есть здесь своя цена и у обоих видов некриптической окраски (Ccol, Cmc) (табл. 1).
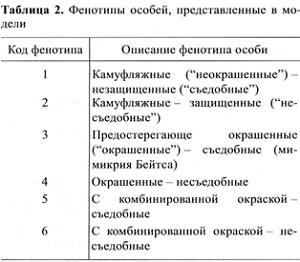
Всего в модели возможны шесть фенотипов особей (табл.2). При этом в любой момент времени распределение особей по фенотипам может быть каким угодно, но общая численность особей всегда равна 1000.
Степень маскировки, выражаемая в вероятности обнаружения особи хищником, задаётся параметром P: вероятность обнаружения криптически или комбинированно окрашенных животных (Рc) ниже, чем вероятность обнаружения их ярко расцвеченных сородичей (Рa) (табл.1).
Под ценой атаки (Cc) подразумевается снижение плодовитости подвергшейся атаке, но не съеденной особи. А вот станет ли хищник атаковать жертву – зависит от скорости его обучения (k) и продолжительности памяти (M) (табл. 1).
Диапазоны параметров – это условно ручки настройки условного осциллографа, покручивая которые, можно получать на экране разнообразные интереснейшие картины.
Придавая параметрам некие правдоподобные значения (или значения, которые кажутся нам правдоподобными) имеем определенный базовый вариант. Модельное решение для него представлено на рис. 2. Мы видим, как на протяжении 1000 поколений колеблется численность особей для каждого из шести фенотипов. Поскольку ни один из фенотипов не вымер (это мы так хитро подобрали базовый набор параметров), то можно моделировать нашу эволюцию хоть на миллион поколений (тактов) – картина существенным образом не изменится.
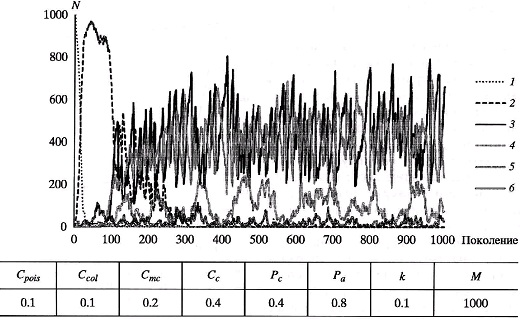
Имея такую замечательную «игрушку», трудно отказать себе в удовольствии подвергнуть мир наших оранжевых мухо-ос различным испытаниям всего лишь лёгким движением ручки настройки какого-нибудь параметра.
Помимо базового сценария проведено еще 8 тестов (по числу ручек настройки). При этом, изменяя один из параметров (ломая базовый вариант), мы всякий раз вкладываем в действие некий биологический содержательный смысл и пытаемся с биологических же позиций осмыслить полученные результаты.
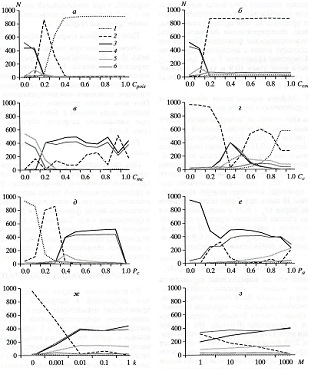
С выбранными базовыми параметрами апосематическая окраска появляется только при небольшой цене ядовитости и некамуфляжной окраски. Однако, например, если качество камуфляжа падает (жертвы перебираются на другой биотоп) или цена камуфляжа перестает быть равной нулю, диапазон параметров существования ярких и комбинировано окрашенных особей расширяется. А вот рост скорости обучения хищников и их хорошая память стимулируют рост численности бейтсовских «обманщиков» — неядовитых особей с предостерегающей окраской (рис. 3, ж, з).
Резюмируя полученные на основе изучения модели результаты, В.И. Грабовский предлагает следующее описание начальных этапов эволюции апосематической окраски:
«1. В силу каких-либо обстоятельств (изменение типа местообитания, смена диеты, мутация окраски и т.д.) камуфляж криптически окрашенных особей ухудшается.
2. Популяционным ответом в этой ситуации может быть выработка жертвами различных средств защиты от хищников (ядов, защитного панциря, иголок, волосков, отталкивающего запаха и т.д.).
3. В результате обучения хищников возникает ассоциативная связь случайных отклонений в окраске жертв с их защищенностью, причем окраска защищенных жертв быстро теряет криптичность и становится заметной для хищника.»
Таким образом, по всей видимости, только что появившаяся апосематическая окраска — это не полезная адаптация, а вредная и неизбежная издержка, результат мутаций, смены среды обитания и т.д. Такие особи становятся более заметны хищникам, и, разумеется, начнётся естесственный отбор на наиболее качественную защиту от хищников с помощью чего-то кроме окраски. Ведь теперь криптическая окраска уже не маскирует особь, и выживать будут те, кто полагался не только на свою окраску, но и на ядовитость, волоски, иголки, чешуйки и т.д. В ответ на это у хищников тоже может начаться отбор на избегание охоты на жертв, которых видно слишком хорошо. Такие жертвы могут быть ядовитыми и колючими, и их есть — себе дороже. В модели это называлось обучаемостью хищников. И, наконец, с этого момента среди жертв станет выгодно иметь апосематическую окраску!
Подобное объяснение ранней эволюции апосематической окраски, конечно, вряд ли можно назвать всеобъемлющим. То есть рискованно было бы утверждать, что апосематическая окраска появилась только так, и точка (хотя, с другой стороны, не исключено, что это правда). Тем не менее, можно вполне уверенно сказать, что подобный эволюционный механизм может играть важную роль в формировании ярких, предостерегающих, окрасок у животных.
Популярные синопсисы