Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Согласованность партнеров укрепляет симбиоз

Координация изменений частот определенных генотипов среди растений и азотфиксирующих бактерий взаимосвязана с экологической эффективностью образованного ими симбиоза. Последняя в свою очередь определяется эпигенетическим наследованием хозяином благоприобретенных симбиотических свойств и альтруизмом бактерий по отношению к своему хозяину. Повышение согласованности партнеров усиливает их интеграцию – вплоть до объединения в новые самостоятельные организмы.
В симбиотическом сожительстве тесное и длительное сосуществование представителей разных биологических видов включает взаимное влияние их биохимических и генетических процессов, и, в конечном счете, определяет их отношение с внешним миром. Несмотря на то, что симбиозы с некоторыми оговорками можно рассматривать как промежуточное звено между взаимодействием и слиянием, связи партнёров становятся столь близкими, а синтезируемые ими друг для друга вещества до такой степени незаменимыми, что происходит становление новой надорганизменной системы. По уровню своей автономности, независимости от окружающей среды и внутреннему единству такая система может приближаться свободноживущему (унитарному) организму, в котором целое главенствует над частями и больше суммы своих частей.
Удобным объектом для изучения взаимосвязи целостности и эффективности симбиотических систем служат азотфиксирующие бактерии ризобии, сосуществующие в союзе с бобовыми растениями (рис. 2). Этой группе симбионтов в основном и посвящена статья Н.А. Проворова и Н.И. Воробьева, сотрудников ГНУ ВНИИСХМ Россельхозакадемии.
В ходе продолжительной – свыше 60 млн. лет – эволюции ризобиального симбиоза усложнение структуры клубеньков (структур особого строения на корнях растений, в которых селятся бактерии) сопровождалось повышением экологической эффективности и усилением генетического контроля над взаимосвязями партнеров.
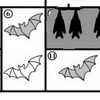
Симбиотические отношения чрезвычайно широко распространены в природе. Уровень целостности в симбиозах возрастает, во-первых, в следующем ряду взаимодействий: факультативные, т.е. необязательные (например, сожительство раков-отшельников с актиниями) →экологически облигатные, обязательные только в определенных условиях (например, энергетически затратная кооперация растений с азотфиксирующими бактериями, возникающая при дефиците азота) → генетически облигатные, обязательный всегда (например, бактерия Carsonella, передавшая во многом полномочия своего генома хозяину-листоблошке (рис. 1), см.: «Прочтён самый маленький геном»). Во-вторых, целостность усиливается при переходе от эктосимбиозов (те же раки-отшельники с актиниями) к эндосимбиозам (например, жгутиконосцы, обитающие в кишечнике термитов и помогающие переваривать целлюлозу). В-третьих, целостность повышается от внутритканевых симбионтов (цианобактерии в межклетниках корня саговников) к внутриклеточным (бактерии, которые как матрешка в матрешке, живут в жгутиконосцах, которые живут в термитах, см. также: «Бактерии, разлагающие для термитов древесину, ещё и разлагают для них атмосферный азот»).

Азотфиксирующие бобово-ризобиальные симбиозы относятся к экологически-облигатным эндосимбиозам (рис. 2). Целостность этих систем возрастает при переходе от симбиозов, в которых бактерии существуют в межклетниках корня к симбиозам внутриклеточным, с образованием в растительных клетках клубеньков специальных структур – симбиосом. Эти структуры состоят из азотофиксирующих бактероидов (бактерий, уже проникших в корень), окруженных мембраной. Бактероиды в симбиосомах могут сохранять сходство со своими свободноживущими родственниками, но могут и резко отличаться от них – большими размерами, повышенным количеством азотфиксирующего фермента нитрогеназы, потерей способности к генетической рекомбинации. Еще одним фактором, повышающим целостность симбиоза, является специализация инфицирования: переход от проникновения бактерий внутрь корня через разрывы его эпидермиса к активному поглощению бактерий самими корневыми волосками.


Экономия углерода, необходимого для синтеза транспортных форм азота, передаваемого из клубеньков в листья растений, и совершенствование строения самих клубеньков также рассматриваются как маркеры специализации и повышения целостности ризобиальных симбиотических систем.
Важнейшим фактором целостности симбиоза является авторегуляция – сигналы, посылаемыми партнерами друг другу на всех этапах развития симбиоза. В самом начале корневые волоски растений привлекают бактерий, находящихся в почве, аттрактантами флавоноидной природы. С флавоноидов начинается сложный межвидовой «диалог» между будущими симбионтами на тему «а подходим ли мы друг другу?» Растение химическим путем оповещает окружающие бактерии о том, что оно ищет себе симбионтов. Рецепторы к флавоноидам, расположенные на мембране бактерий, восприняв сигнал, изменяют активность транскрипционных факторов, влияющих на работу генов. В результате запускатся бактериальные гены, ответственные за синтез липоолигосахаридных молекул - так называемых Nod-факторов. Эти молекулы, в свою очередь, являются «ответом» бактерии растению в этом диалоге. Nod-факторы специфичны для разных штаммов ризобиев. Следовательно, «разговор» о симбиозе продожается только в том случае, если выделямый бактерией Nod-фактор свидетельствует о том, что бактерия растению подходит. То есть в случае будущей эффективности симбиоза. Кроме Nod-факторов, со стороны растений «нужные» им бактерии распознаются по полисахаридам. Восприятие растением ответа бактерий происходит с участием белков лектинов (см. также: «Лектины заменяют растениям иммунную систему»). На более поздних стадиях пептидный комплекс CLAVATA и другие сигнальные факторы, синтезируемые растениями, продолжают контролировать образование клубеньков.
В общем, обмен сигналами у симбионтов выходит за пределы обмена источниками питания. Складывается надорганизменная система «симбиогенома», «с его эволюцией от функциональной интегрированности партнеров (…) к структурному объединению их генных систем». Авторы приводят множество примеров достижения целостности межвидовых взаимодействий: паразитических высокоспециализированных грибов с растениями, мутуалистических, т.е. взаимовыгодных союзов растений с азотфиксирующими цианобактериями, растительноядных насекомых с «пищеварительными» бактериями, лишайников, скомпонованных грибами и цианобактериями (или грибами и зелеными водорослями, или всеми троими вместе) (см. также: «От биохимического сотрудничества – к общему геному»). Иногда специализация и взаимная интеграция организмов столь высока, что родители «вертикально» передают своих симбионтов потомству (см.: «Клопы кормят своё потомство полезными бактериями»).
Совместная эволюция партнеров, коэволюция, рассматривается как процесс, выражающийся в «тяготении» симбиоза «к организменному уровню организации» вплоть до перехода к единому организму. Способствовать такому переходу может как миграция партнеров в новые экологические зоны, поддерживающая их единство, так и наследование симбионтов хозяином (вертикальная передача), зачастую вызывающая утерю симбионтами частей генома.
В статье приведено краткое описание математических моделей, разработанных ранее (см.: Н. И. Воробьев, Н. А. Проворов, «Моделирование эволюции бобово-ризобиального симбиоза при мультиштаммовой конкуренции бактерий за инокуляцию симбиотических ниш», «Моделирование эволюции бобово-ризобиального симбиоза на повышение функциональной интегрированности партнёров и экологической эффективности их взаимодействия»). Эти модели показывают, что возрастание функциональной интеграции партнеров бобово-ризобиального симбиоза (в смысле согласованного изменения частот генотипов партнеров под влиянием внешних факторов) связано с симбиотической эффективностью (продуктивностью партнеров). В данном симбиозе сожители все еще сохраняют автономность. Для поддержания целостности здесь важно такое качество как межвидовой альтруизм – наличие свойств бесполезных или даже вредных для самого их носителя, но зато ценных для партнера. В случае клубеньковых симбионтов это превращение ризобия в бактероид – продуктивно фиксирующий азот во благо растения, но потерявший способность размножаться. Впрочем, бактерии восполняют свою утрату благодаря углероду, которым растение щедро делится с ними. Таким образом, растения и бактерии здесь связаны положительными обратными связями (потоком азота в растение, создаваемым бактериальной клеткой – чем больше этот поток, тем выше интенсивность фотосинтеза и тем больше его углеродистых продуктов растение передает клубенькам). Растения, как оказалось, способны не пускать в свои клубеньки «неподходящих» для себя неактивных бактерий-азотфиксаторов, предоставляя преимущество специфичным для себя активным штаммам (фиксирующим азот только с данным растительным генотипом). Именно в таких специфичных сочетаниях эффективность симбиоза была максимальной, что создавало предпосылки для эволюционного закрепления удачных сочетаний, обеспечивающих повышение целостности симбиосистемы.
Авторы статьи формализуют этапы эволюции целостности симбиозов. Первый этап - координация метаболизма и регуляторных функций в комбинациях согласованных генотипов. Затем, после дальнейшей координации биохимических реакций, закрепленных в тканевых и клеточных структурах партнеров, приходит время для следующего важного этапа интеграции. Он состоит в вертикальном наследовании симбионтов хозяином и сопряжении генетической информации партнеров благодаря горизонтальному переносу генов (см. также: «Животные обмениваются генами с паразитическими бактериями», «Горизонтальный обмен генами заменяет коловраткам половое размножение», «Бактериальные гены помогают нематодам паразитировать на растениях»). Завершающим и необратимым этапом является упрощение генотипов симбионтов, как это произошло при формировании эукариотической клетки из бактерий и архей (см. также: «Новый сценарий происхождения эукариот», «Фармальные статистические тесты подтвеждают происхождение всех живых организмов от единого предка»; «Для эукариот более важны гены, полученные от архебактерий»).
В статье затронут вопрос эпигенетического наследования признаков симбиоза, который более полно освещается в одной из предыдущих работ этих же ученых. Наличие эпигенетических программ проявляется в том, полагают авторы, что совместными усилиями симбионты создают новые органы – клубеньки, в то время как порознь ни один из них не имеет полного набора генов для этих инноваций. Регуляторные растительные гены контролируют работу как собственных, так и бактериальных симбиотических генов, бактериальные гены также могут быть регуляторами. Наблюдается функциональная интеграция геномов обоих симбионтов – симбиогенез, т.е. заново возникают не только морфологические структуры клубеньков, но и сама система наследственности. Причем примерно тот же набор генов растений, который отвечает за ризобиальный симбиоз, может быть задействован в иных условиях в другой программе – формировании микоризы (см. также: «Симбиоз – основа растительной жизни»). Таким образом клетка, «получая различные сигналы, обладает способностью выбора программы, в которой ей, а точнее ее генам, наиболее целесообразно участвовать». Эволюция различных симбиозов показывает, что с течением времени адаптивные симбиотические признаки могут наследоваться – через вертикальную передачу хозяином симбионта, а это значит, что хозяин имеет «возможность направленно приобретать полезные симбиотические свойства вместе с их носителями – микроорганизмами». Вероятно, из этого можно заключить, что мы имеем дело с адаптивными изменениями функционального состояния симбиогенома, предваряющими его жестко наследуемую генетическую фиксацию, т.е. с наследованием благоприобретенных признаков (см. также: «Развитие эволюционной теории невозможно без молекулярной генетики»; страничка про эпигенетическую теорию эволюции).
Авторы считают, что симбиогенез богаче эволюционными возможностями, нежели эпигенез унитарного организма, потому что симбиоз объединяет разнородные генотипы прокариот и эукариот, в то время как единый организм ограничен экспрессией одного и того же генотипа в различных тканях и клетках.
Рассмотрев симбиотические стратегии у насекомых, позвоночных животных и растений, авторы приходят к выводу, что растения в большей степени, чем многоклеточные животные, сохранили в ходе эволюции способность к существованию в форме стабильных симбиотических систем.
Изучение разных типов симбиозов позволяет сопоставить структурно-функциональную целостность, основанную на координированных изменениях популяций партнеров, с эффективностью этих популяций и исследовать многообразные пути процесса эволюции органических форм.
Популярные синопсисы