Последние выпуски
- Том 87, № 2. Март-апрель 2026
- Том 87, № 1. Январь-февраль 2026
- Том 86, № 6. Ноябрь-декабрь 2025
- Том 86, № 5. Сентябрь-октябрь 2025
Экология для статистики – целина или пустыня?

Многочисленные статистические методы более или менее успешно используются в экологических исследованиях при анализе изменений пространственной структуры сообществ. Эти методы позволяют сравнивать эмпирические данные с «нулевыми гипотезами», постулирующими отсутствие связи между изучаемыми переменными. Это необходимо для того, чтобы ответить на главный вопрос, мучающий любого исследователя: «А не случайность ли полученный результат?» Однако адекватность даже современных статистических заключений сильно зависит от массы факторов, которые необходимо учитывать при каждом исследовании. К этим факторам можно отнести степень детализации исходных данных, масштаб и уровень группировки блоков исследуемых экосистем. Предложенные методы позволяют в общем-то эффективно определять достоверность полученных результатов, т.е., в случае изучения экологических сообществ, статистические методы лишь сигнализируют о наличии взаимосвязи между видами. Однако само объяснение взаимосвязи: межвидовыми взаимодействиями, влиянием окружающей среды и т.д. – по-прежнему остается уделом специалистов-экологов. В целом, в статье решается вопрос о корректности применения ряда методик (например, анализа «вложенности- nestedness») нового, пока ещё для многих непривычного, уровня.
Компьютерный прогресс позволяет решать задачи, которые еще лет 20 назад даже не поднимались из-за отпугивающей сложности математического аппарата и громоздкости вычислений. Образно говоря, когда компьютеры были большими, а их память и быстродействие – незначительными, время вычислений для иных реальных задач составляло бы годы, а то и столетия. Вот и пылились, «задвинутые на полку», идеи и алгоритмы, опередившие время (например, работа Хюлтена по оценке упорядоченности таксонов сообществ, увидевшая свет еще в 1937 г.). Но время пришло.
Целью исследования сотрудников Института экологии Волжского бассейна В.К. Шитикова и Т.Д. Зинченко была проверка работоспособности различных современных методик оценки организации видовой структуры сообществ на примере макрозообентоса – мельчайших, но различимых глазом водных придонных животных.

Исследуемая речная система типична для малых рек бассейна средне-нижней Волги с весьма разнообразными биотопами и ландшафтными элементами. За период 1990-2007 гг. было взято 147 гидробиологических проб, в исходные матрицы для статистического анализа включено 375 видов и других таксономических групп зообентоса.
Структура сообществ может объясняться разнообразием условий их обитания; также структура может быть следствием межвидовых отношений: конкуренции, хищничества, паразитизма, мутуализма и т.д. Эти отношения определяют динамику структурных преобразований сообществ. По мнению ряда западных ученых, эта динамика контролируется тремя основными процессами: становление взаимно согласованных отношений между видами, их взаимное исключение и формирование обособленных групп. Реализация этих процессов в преобразовании сообществ приводит к одному из 4-х типов пространственной структурны: стохастичной (рис. 3, а), упорядоченной (рис. 3, б), с взаимозаменяемостью видов (рис. 3, в) или с образованием специфичных локальных ассоциаций видов (рис. 3, г).

Мерой упорядоченности структуры является т.н. «вложенность» (nestedness). Усилиями многих авторов (корни, как раз, в упомянутом 1937 г., но особо отметим Атмара и Паттерсона в 90-х годах) nestedness-анализ стал основным экологическим инструментом исследований по данной тематике.
Более подробное описание понятия вложенности можно найти в монографии «Макроэкология речных сообществ: концепции, методы, модели», опубликованной авторами обсуждаемой статьи ранее. Представим себе, например, острова, расположенные на различных расстояниях от материка. Если на близких к материку островах проживает полный комплект материковых видов, а по мере удаления от берега видовое богатство островов убывает, то мы имеем дело с высокой вложенностью объекта. Т.е. видовой состав дальних островов является заранее предопределенной частью состава ближних островов (рис. 3, б). Если же виды на островах распространены приблизительно равномерно, и островные сообщества являются просто случайным фрагментом материковой флоры/фауны, то существенной организованности мы не заметим (рис. 3, а).
Исследуемую речную систему разбили на 3 участка согласно гидрологическим характеристикам. Для каждого участка и всего водотока в целом составили матрицы (таблицы) размером s x m, где s- число видов, m - количество проб (или местообитаний, т.е. объединенных проб, соответствующих отдельным биотопам). Далее эти матрицы сортировали так, чтобы в верхних строках оказывались виды (единицы) с наибольшей встречаемостью, в столбцах слева – местообитания с наибольшим видовым богатством (рис. 4). Выраженность эффекта вложенности оценивали как концентрацию «единиц» в верхнем левом треугольнике отсортированной матрицы и концентрацию «нулей» в нижнем правом углу.
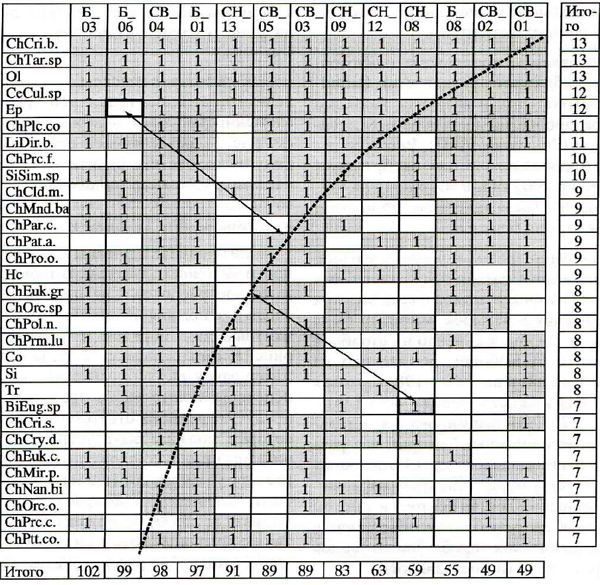
Весьма образной является такая единица измерения вложенности как температура, Т (разумеется, к физической температуре (градуснику) это не имеет никакого отношения). Подобно температуре жидкого состояния воды Т меняется от 0о до 100о. Нулевая температура - идеальная вложенность - соответствует рис.3.б; приближение к 100о определяет случайный тип организации сообщества – рис.3а. Другими словами, вот – 0о, устойчивая, консервативная структура сообщества, буквально скованная льдом устоявшихся связей, а вот – 100о, кипение, хаос, самые немыслимые связи (или их отсутствие) между таксонами сообщества. Непосвященным может показаться странной беседа специалистов, дескать, почему же температура Волги в Астрахани на 30о выше, чем в Самаре… Температура вложенности, Т – это суммарное расстояние всех «единиц» и «нулей», оказавшихся не в «своей» зоне, до диагонали, зоны эти разделяющей. Вложенность рассчитывали еще несколькими способами: в том числе, с помощью BR-индекса (количество «неуместных» единиц и нулей), NODF-индекса (число взаимосвязанных пар видов или участков).
Полученные результаты сравнивали с пятью нуль-моделями. Нуль-модели подразумевают заведомое отсутствие взаимосвязи между изучаемыми переменными. Чтобы добиться этого в данном случае, исходные виды и местообитания были случайно перемешаны (рандомизированы) разными способами. Если эмпирические значения переменных достоверно отличались от аналогичных, но полученных в результате перемешивания, то считалось, что в природных сообществах наблюдается упорядоченность.
Оказалось, что вложенность, выраженная через Т- и BR-индексы, зависела от размера обработанных данных и от детализации, т.е. от того, объединялись ли пробы в группы: отдельные пробы и объединенные по биотопам пробы давали разный результат, в то время как NODF-индекс, также зависимый от детализации, все же показал совпадающий вывод как для отдельных, так и для агрегированных проб.
Ученые отмечают, что рассмотренные индексы не указывают на отличие типов биоты (например, образовано ли сообщество речными или озерными видами), они близки между собой для всех трех исследованных водотоков, несмотря на своеобразие видов каждого из них.
Показатели Т- и BR-индексов, рассчитанные для отдельных видов, также неоднозначно характеризуют их экологию. Так, высокие индексы могут быть как у относительно распространенных видов, которые четко приурочены к локальным особенностям среды, так и у чужеродных случайных мигрантов из соседних зон.
Индексы, рассчитанные для местообитаний, показали высокие «температуры» верхней части р. Байтуган и устья р. Сок, что свидетельствует о специфическом видовом составе этих участков. Такое выделение «особенных» местообитаний важно для планирования охраняемых территорий.
Для того чтобы понять, насколько часто согласованность видов (одновременная встречаемость пары видов) может оказаться случайной, строки и столбцы исходных матриц также многократно перемешивались. Для каждой пары видов сравнивали согласованность (С) исходной матрицы со средним значением С перемешанных матриц, и отбирали (отфильтровывали) только те комбинации пар, которые достоверно отличались от ожидаемого случайного С.
Характер межвидовых взаимодействий для всей экосистемы в целом оценивали с помощью V-отношения Шлютера: отношения общей дисперсии обилия видов к сумме дисперсий для каждого из них. При отсутствии таких отношений (нуль-гипотезе) V=1, а значимо большее или меньшее единицы V указывало на положительное или отрицательное взаимодействие между видами в сообществе.
Приведенные в статье статистические фильтры позволили пересчитать степень доверия к полученным данным, сократить количество анализируемого материала и избежать издержек гипердиагностики. Однако выявленные математически положительные и отрицательные связи между таксонами нуждаются в биологическом толковании. Так, взаимное исключение видов может объясняться и неоднородностью условий среды, и конкуренцией видов на сходных биотопах. Согласованная встречаемость пар видов определяется, вероятно, сходством их экологических пристрастий: например, повышенной требовательностью к наличию кислорода в воде или обоюдным тяготением к заиленному дну. V-отношение Шлютера оказалось меньше единицы, что может обуславливаться как межвидовыми отношениями (хищничество, конкуренция), так и различиями обилия ресурсов, приводящими к колебаниям обилия видов. Авторы отмечают, что найденные закономерности лишь сигнализируют о наличии взаимосвязи между видами, но поиски смысловой подоплеки – задача специалистов-экологов.
Во многих случаях, полученные в работе выводы можно было бы считать тривиальными (или, мягче говоря, «консервативными»), но задачи авторов лежат, скорее, в методологической сфере.
Недостатки описанных методов в полной мере присущи и старым количественным индексам (например, определения индекса сходства по Съеренсену - отношение числа общих для сравниваемых биоценозах видов к сумме видов, обнаруженных в каждом этих биоценозов), а вот достоинства налицо – расширение спектра параметров оценки видовой структуры обеспечит несомненный прогресс при изучении экологии сообществ. Лавинообразная компьютеризация, по большому счету, делает практически идентичными по трудозатратам расчет температуры вложенности, V-отношения Шлютера или традиционных индексов.
Популярные синопсисы