Новая легенда о Кракене

В конце апреля в высокорейтинговом журнале Science вышла статья “Earliest octopuses were giant top predators in Cretaceous oceans” («Первые осьминоги были гигантскими хищниками высшего уровня в океанах мелового периода»), подготовленная коллективом авторов, большинство из которых работают в различных университетах Японии. На основании изучения изолированных находок крупных челюстей головоногих моллюсков они пришли к выводу, что в морях мелового периода процветали гигантские осьминоги, настоящие «кракены», которые, как минимум, конкурировали с гигантскими позвоночными хищниками за добычу, а может быть даже и охотились на них. Оценки максимальной длины этих осьминогов варьируются в очень широком диапазоне — от 7 до 19 метров, но именно последнему значению в статье, на иллюстрациях к ней и уж конечно во всех пресс-релизах уделено гораздо больше внимания, чем всем остальным вариантам оценки. Да и от названия статьи веет уверенностью: «осьминоги были гигантскими хищниками», а не «могли быть», как часто пишут более осторожные палеонтологи. А на самом деле, есть ли основания для такой уверенности?
Исследование, результаты которого опубликованы в Science, базируется на изучении изолированных нижних челюстей головоногих моллюсков. Среди них как описанные ранее другими палеонтологами образцы (см. картинку дня «Гигантские челюсти мелового моллюска»), так и собственные находки авторов новой работы. Челюсти происходят из верхнемеловых отложений Японии и Канады (остров Ванкувер) и имеют возраст от 100 до 72 млн лет. Этот интервал охватывает почти весь верхний мел — с начала сеномана по конец кампана. Исходя из формы челюстей, авторы новой публикации сделали вывод, что они принадлежали цирратным осьминогам (подотряд Cirrata).
В настоящее время цирратные (они же плавниковые) осьминоги обитают на больших глубинах, но в мезозое, возможно, жили и в мелководных морях и на окраинах океанов. На руках у них присутствуют не только присоски, но и усики-цирры (отсюда и их название). Наличие этих усиков, пара плавников, а также не полностью редуцированная внутренняя пластинка — гладиус, роднят этих осьминогов с их предками — вампироморфами, и большинство исследователей полагает, что древнейшие осьминоги должны были относиться именно к подотряду Cirrata. Но ни гладиусов (представляющих собой по сути редуцированный до тонкой органической пластинки рудимент внутренней раковины), ни тем более отпечатков мягких тканей цирратных осьминогов в ископаемой летописи пока не найдено, хотя такие находки известны для второго подотряда осьминогов — Incirrata.
В распоряжении авторов новой статьи тоже были не гладиусы, а только отдельные челюсти. Стоит отметить, что среди моллюсков только головоногие — уникальные обладатели хорошо развитого челюстного аппарата. У них есть пара челюстей — верхняя и нижняя, между которыми расположена радула — подвижный «язык», покрытый рядами острых зубчиков. Радула присутствует у всех моллюсков, кроме двустворчатых, а вот верхняя и нижняя челюсти — это эволюционное приобретение головоногих. Челюсти снабжены мощной мускулатурой и вместе с радулой заключены в своеобразный мускульный мешок: спереди из него выступают только передние концы челюстей, сзади он открывается в пищевод. Друг с другом челюсти соединены посредством мышц и в ископаемом состоянии сохраняются, как правило, по отдельности.

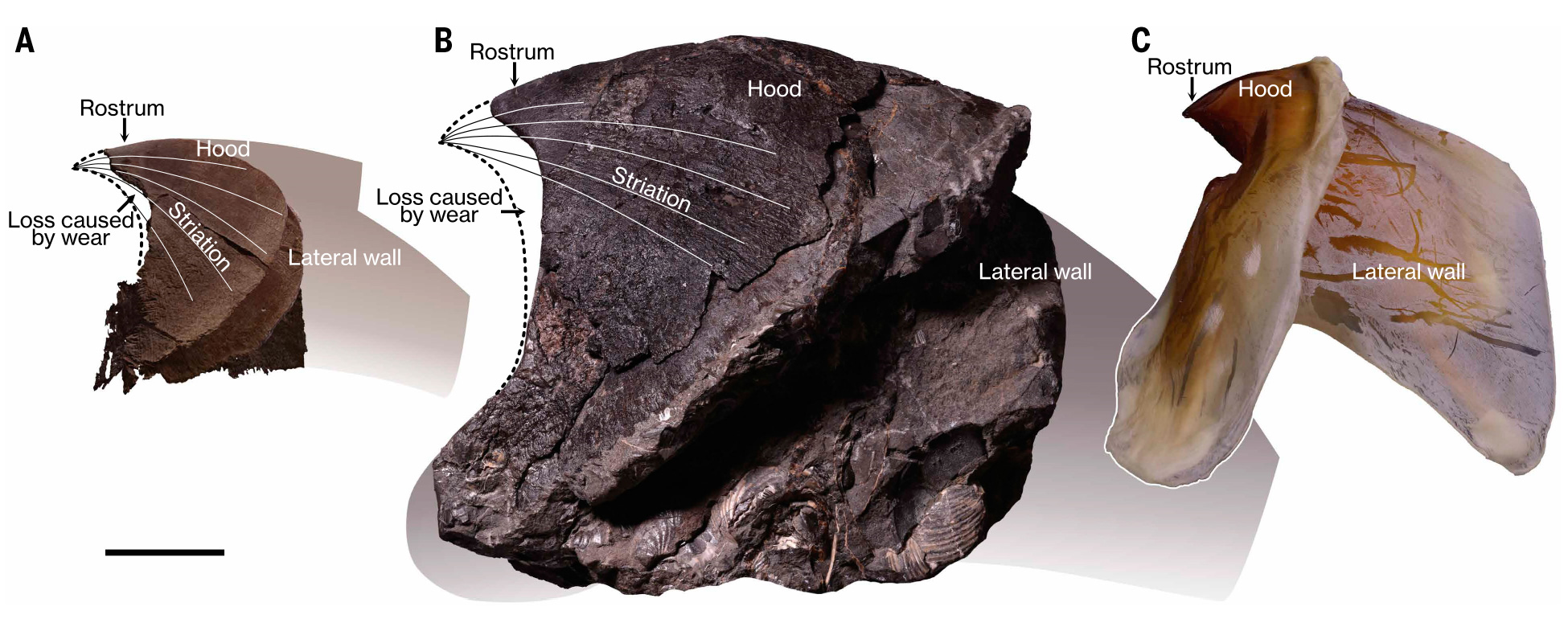
Строение челюстей колеоидеи на примере каракатицы Sepia ofcinalis. А — схема строения верхней челюсти (hood — это капюшон, rostral tip — острие челюсти, crest и lateral wall образуют рукоятку), В — схема строения нижней челюсти (hood — капюшон, wings — крылья, боковые выросты капюшона, lateral wall и crest — рукоятка). C и D — стандартные измерения челюстей. В случае ископаемых находок наибольшее значение имеет длина капюшона (UHL и LHL в верхней и нижней челюстях соответственно). Общую длину (UTL и LTL) обычно очень сложно измерить из-за неполной сохранности рукоятки. Иллюстрация из статьи L. Souquet et al., 2023. The growth of the buccal mass in Sepia officinalis: functional changes throughout ontogeny
Из-за сходства с клювами некоторых птиц (в первую очередь попугаев) челюстной аппарат головоногих нередко называют клювом, причем этот термин используется и в научных работах. С помощью челюстей головоногие могут быстро убивать добычу и откусывать от нее большие куски, которые затем перетираются при помощи радулы. Каждая челюсть состоит из двух пластин — наружной и внутренней; наружная называется капюшоном, внутренняя — рукояткой. Челюсти состоят из органического вещества — хитина. У современных наутилусов (начиная с середины триаса) и у многих вымерших аммоноидей челюсти дополнительно укреплены кальцитовыми элементами. Но в подклассе Coleoidea, к которому относятся осьминоги, кальмары и другие головоногие, не имеющие наружной раковины, кальцитовые элементы в челюстях никогда не возникали. Тем не менее, их челюсти, изначально полностью органические, в принципе неплохо сохраняются в ископаемом состоянии за счет замещения различными минералами (в основном апатитом в ходе процесса фосфатизации). Чаще всего бывает, что именно клювы, причем распавшиеся (верхняя и нижняя челюсть по отдельности) — это всё, что сохраняется в палеонтологической летописи от древнего головоногого моллюска.

Сравнение челюстей осьминогов и кальмаров. Сверху верхняя челюсть, снизу нижняя. 1 — инцирратный осьминог Muusoctopus, 2 — цирратный осминог Opistoteuthis, 3 — кальмар Taningia, 4 — кальмар Todarodes. Из статьи J. C. Xavier, Y. Cherel, 2021. Cephalopod Beak Guide For the Southern Ocean: an update on taxonomy
Хотя у всех головоногих челюсти устроены в принципе одинаково, в разных эволюционных линиях наблюдаются существенные различия в форме и пропорциях различных элементов челюстей. Например, передний конец верхней челюсти у кальмаров и каракатиц всегда заметно длиннее и острее, чем у осьминогов, зато у осьминогов рукоятка нижней челюсти всегда заметно длиннее, чем у вампиротеутид — их вероятных предков. Эти различия позволяют по морфологии челюстей современных головоногих определять, кому именно они принадлежали. С ископаемым челюстями сложнее: их форма может быть существенно искажена до или во время фоссилизации. Но об этом чуть позже.
Nanaimoteuthis jeletzkyi, нижние челюсти с многочисленными следами разложения и неполной фоссилизации, преимущественно в задней (наиболее тонкой) части образцов. Дополнительные материалы к обсуждаемой статье
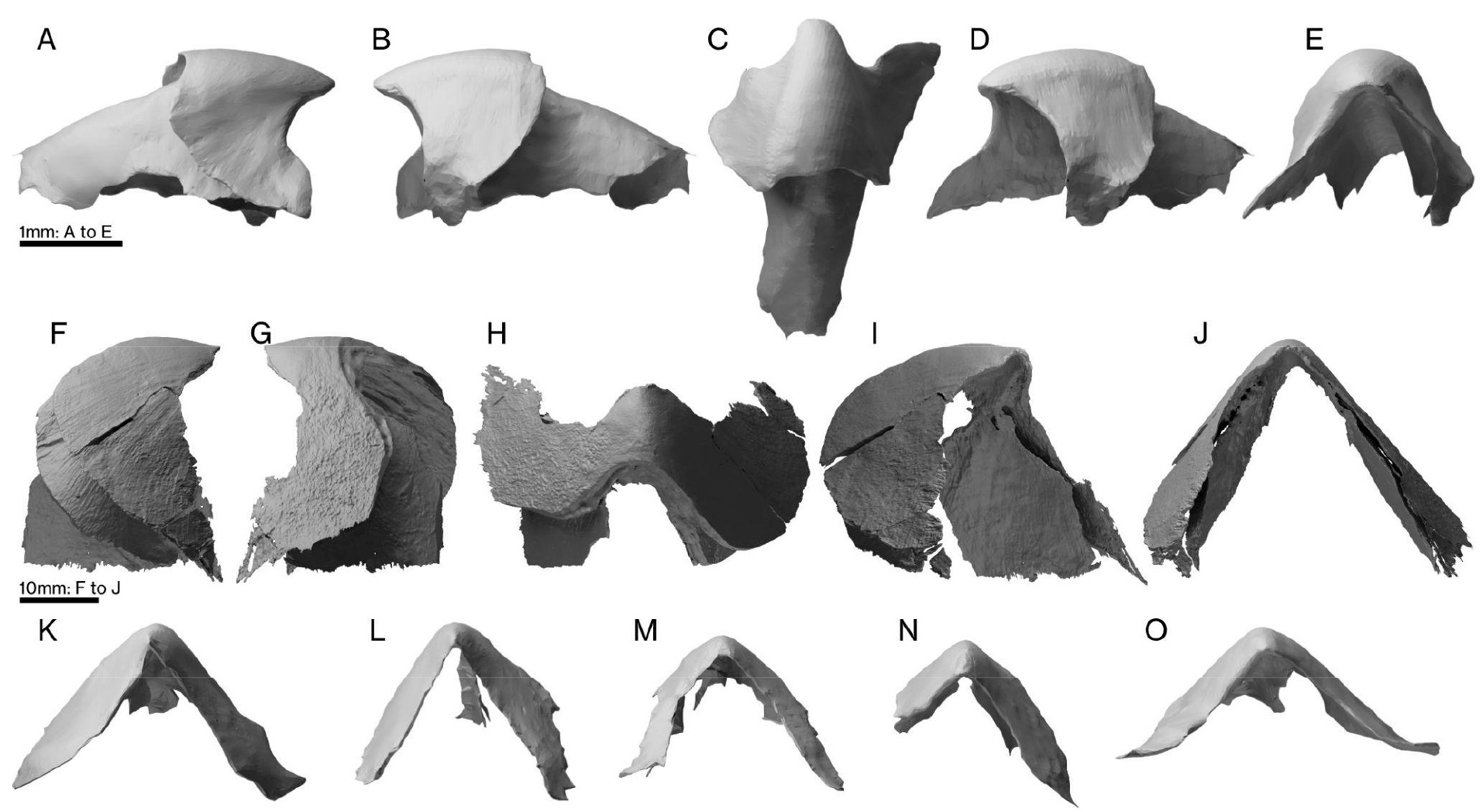
Именно возможность по морфологии челюстей вычислять их обладателей легла в основу нового исследования. В распоряжении авторов были 15 ранее найденных (см. картинку дня «Гигантские челюсти мелового моллюска») крупных нижних челюстей ископаемых головоногих (дополнительные материалы к статье, table S1) а также 12 нижних челюстей из собственных материалов, имевших более скромные размеры (дополнительные материалы к статье, table S2). Верхние челюсти авторы включать в эту подборку не стали, сославшись на то, что в них меньше важных морфологических признаков. Основное внимание исследователи сосредоточили на двух крупных экземплярах. Эти нижние челюсти, отнесенные авторами новой публикации к видам Nanaimoteuthis jeletzkyi и N. haggarti, имели длину наружной пластины (капюшона) 39,6 мм и 86,4 мм соответственно (дополнительные материалы к статье, table S4).
Ископаемые челюсти Nanaimoteuthis. A — N. jeletzkyi, B — N. haggarti, C — нижняя челюсть современного гигантского кальмара Architeuthis dux. Масштабный отрезок 20 мм. Пунктиром и серым цветом показаны не сохранившиеся, по мнению авторов, части челюстей. Рисунок из обсуждаемой статьи.
Здесь нужно уточнить, что род Nanaimoteuthis, описанный несколько лет назад (K. Tanabe et al., 2016. Late Cretaceous octobrachiate coleoid lower jaws from the north Pacific regions) — это не род головоногих моллюсков как таковой, а паратаксон, выделенный по изолированным челюстям. Но для того, чтобы не громоздить каждый раз пояснения в стиле «обладатели челюстного аппарата, отнесенного к Nanaimoteuthis», я буду использовать это название и для обозначения самих владельцев этих челюстей. Интересно, что первоначально Nanaimoteuthis был отнесен к отряду Vampyromorpha, к предкам осьминогов, из-за близкого сходства ископаемых челюстей с челюстями других вампироморф. Авторы нового исследования перенесли его в подотряд цирратных осьминогов. На том, чем именно они руководствовались в принятии такого решения, я позже остановлюсь подробнее.

Разные виды цирратных осьминогов: видно разнообразие их морфотипов. Рисунок из книги P. Jereb et al., ed., 2016. Cephalopods of the World. Volume 3. Octopods and Vampire Squids
К сожалению, о том, как выглядели обладатели челюстей Nanaimoteuthis, не известно ничего. Среди цирратных осьминогов, к которым авторы отнесли нанаймотеутисов, существует большое многообразие форм: с коротким телом и с вытянутым телом, с короткими руками, едва превышающими длину тела, и с длинными — в несколько раз длиннее мантии. Для каждого вида существует определенное соотношение длины челюстей и мантии (дополнительные материалы к статье, table S3). Этим и воспользовались исследователи. Они взяли данные по современным осьминогам (кстати, гигантов среди них не оказалось, самая большая челюсть имела длину 13 мм), а затем для каждого вида в имеющееся соотношение подставили длину самых крупных ископаемых челюстей Nanaimoteuthis.
Правда, перед этим они условно разделили осьминогов на обладающих коротким телом и длинным телом, и постулировали, что Nanaimoteuthis должен был быть длиннотелым. Обосновано это было двумя аргументами. Во-первых, согласно молекулярно-генетическим данным, ныне живущие рода с длинным телом возникли раньше, чем короткотелые. Это действительно так (M. Taite et al., 2023. Genome skimming elucidates the evolutionary history of Octopoda), но с учетом того, что речь идет о позднемеловой эпохе, когда уже вовсю процветали и значительно более молодые инцирратные осьминоги (см. картинку дня Ископаемые осьминоги), эта логика выглядит странной: те, древние осьминоги, жили гораздо раньше. Во-вторых, авторы указали, что в морфологии челюстей широкие крылья характерны для длиннотелых форм, тогда как у короткотелых форм крылья узкие. Но никаких ссылок, подтверждающих это утверждение, они не привели.
Однако, даже без учета отброшенной части вариантов, результаты расчетов показали огромный разброс значений. Так, если бы древние осьминоги имели пропорции современного Cirroteuthis muelleri, то обладатели клювов Nanaimoteuthis jeletzkyi имели бы длину мантии всего 0,67 метра, а Nanaimoteuthis haggarti — 1,58 метра. Обладай они пропорциями современного Cirroctopus glacialis, длина их мантии составляла бы 0,76 м и
1,65 м, соответственно. Мелковато для Кракена! Зато пропорции Grimpoteuthis sp., пойманного у южной оконечности Австралии, оказались куда более интересными: для N. jeletzkyi они дают длину мантии 1,84 метра, а для N. haggarti — и вовсе 4,24 метра. Не подвел и Stauroteuthis syrtensis — с его пропорциями N. jeletzkyi получился чуть поменьше — всего 1,64 метра, зато N. haggarti «подрос» аж до 4,43 метра (дополнительные материалы к статье, table S4). Возможность того, что пропорции древних осьминогов, сто миллионов лет назад живших в исчезнувших ныне мелководных экосистемах, могли вообще не укладываться в рамки пропорций современных глубоководных видов, авторы решили не учитывать.
Но мантия — это не всё тело осьминога: есть еще руки, пропорции которых также очень сильно различаются у разных современных видов. Авторы исследования привели данные по 12 таксонам современных цирратных осьминогов, для которых были измерены длина мантии и общая длина с учетом мантии и рук, а из этих измерений рассчитано отношение общей длины к длине мантии. Как и ожидалось, оно тоже варьировало в широких пределах: от 2,7 аж до 7,7. Исследователи взяли среднее значение: 4,2. Не медианное, как чаще делают в таких случаях, а именно среднее — видимо потому, что оно больше. На основании этого отношения N. jeletzkyi с длиной мантии от 0,67 до 1,84 метра получал общую длину от 2,8 до 7,7 м, а N. haggarti с его минимальной длиной мантии 1,58 и максимальной 4,43 метра оказывался в диапазоне от 6,6 до 18,6 метра! Небольшое округление — и в мезозойских морях появляется Кракен длиной 19 метров — длиннее любой морской рептилии, жившей по соседству.

Соотношение длины мантии осьминога по вертикальной оси и длины капюшона (наружной пластины) нижней челюсти по горизонтальной оси, разброс результатов для Nanaimoteuthis и наложенные на вертикальную ось изображения различных морских хищников, убеждающие читателя в грандиозности размеров нового «кракена». Рисунок из обсуждаемой статьи.
По мнению авторов статьи, этот гигант как минимум успешно конкурировал с позвоночными хищниками, такими как акулы и морские рептилии, а может быть даже, будучи высшим хищником морских экосистем, и закусывал своими зубастыми конкурентами. По крайней мере, исследователи нашли на челюстях следы износа, свидетельствующие, по их мнению, о том, что при жизни этот моллюск разгрызал твердые раковины и даже кости.
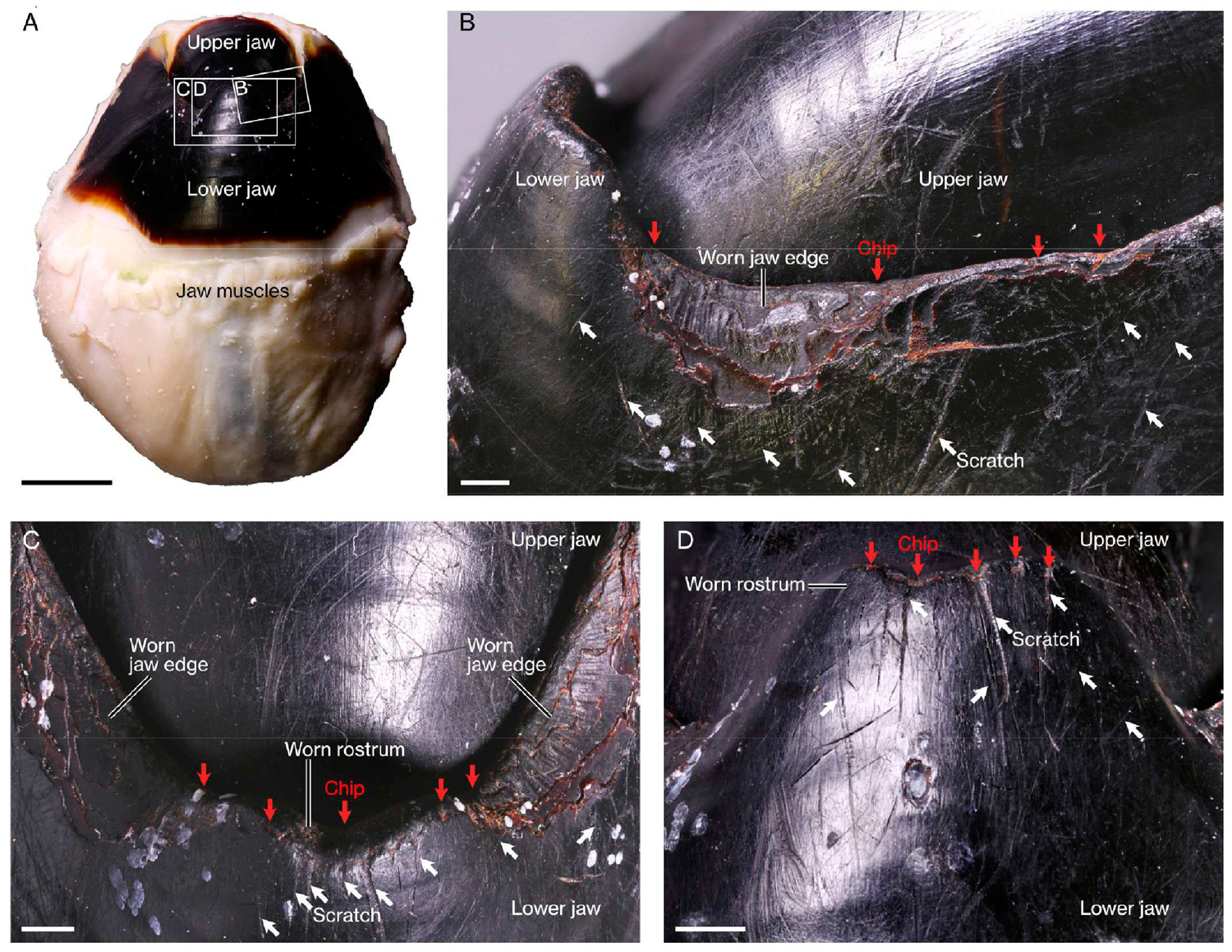
Следы истертости на клюве у современного инцирратного осьминога Enteroctopus. В отличие от современных цирратных осьминогов, питающихся планктонными ракообразными, инцирратные осьминоги включают в свое меню крупных крабов и лангустов. Дополнительные материалы к обсуждаемой статье
Эти же следы износа авторы считают свидетельством того, что он был очень умным. Дело в том, что царапины и следы стесывания на челюсти расположены справа и слева неравномерно, и это дало основание исследователям сделать вывод о том, что у этих «кракенов» имела место латерализация мозга — явление, когда разные полушария мозга берут на себя разные функции, и животное становится правшой или левшой. Считается, что для появления латерализации мозг должен быть очень хорошо развит. Впрочем, у современных осьминогов латерализация известна (R. A. Byrne et al., 2002. Lateral asymmetry of eye use in Octopus vulgaris), и у их мезозойских предков она тоже вполне могла быть.

Царапины в центральной части N. Jeletzkyi — вероятные следы прижизненного износа при разламывании раковин и панцирей добычи. Фото из обсуждаемой статьи.
Гораздо сложнее и важнее ситуация с износом челюсти, или с тем, что авторы за него принимают. Отдельные царапины и сколы, фото которых приведены в статье, вопросов не вызывают: они бывают и на челюстях современных головоногих, которые питаются лангустами, крабами и двустворчатыми моллюсками. Для того, чтобы их получить, совсем не обязательно грызть кости мозазавров. Но в случае с Nanaimoteuthis авторы полагают, что голотип N. haggarti спереди был стесан почти на сантиметр! Насколько мне известно, такой износ у современных головоногих не задокументирован. Свой вывод авторы сделали, проследив за миниатюрными бороздками на поверхности челюсти и предположив, что они должны сходиться к ее острию (они также хорошо видны на главной иллюстрации картинки дня «Гигантские челюсти мелового моллюска» на левом верхнем снимке). Но эта особенность строения челюстей пока еще плохо изучена: у современных клювов таких бороздок не видно, скорее всего они проявляются на определенной стадии разрушения челюстной стенки. Сходятся ли они именно к острию, либо просто к переднему краю — вопрос, который требует отдельного изучения.
Хотя авторы всё объясняют прижизненным стачиванием, хорошо видно, что челюсть N. haggarti имеет посмертную деформацию — она сильно приплюснута с правой стороны от острия. Это не удивительно: не имеющие минерального покрытия челюсти колеоидей довольно эластичные, после смерти моллюска они часто деформируются при захоронении.

Nanaimoteuthis haggarti в трех проекциях; на центральном снимке видна посмертная деформация образца (отмечено стрелочкой), его правая сторона более пологая, острие челюсти также немного смещено вправо. Такие деформации — частое явление среди ископаемых челюстей головоногих моллюсков, которые до фоссилизации были довольно эластичными. Фото с сайта nationalgeographic.com
Кроме того, существует и проблема неполной минерализации. Изначально челюсть состоит из органического вещества — хитина, но в ископаемом состоянии он не сохраняется, и мы находим только те челюсти, в которых хитин замещен каким-либо минералом (чаще всего вариациями апатита). Но для того, чтобы процесс минерализации начался, челюсть должна оказаться в определенных физико-химических условиях в донном осадке. Попасть туда она может почти сразу после смерти моллюска, а может и существенно позже, когда челюсть уже подверглась разложению, иногда довольно значительному. Все это очень усложняет определение таксономической принадлежности ископаемых челюстей, конечно если подходить к этому делу ответственно. А в изученных челюстях Nanaimoteuthis явно имели место и посмертная деформация, и растворение (по крайней мере задних частей). В этом плане особенно показательна челюсть N. Jeletzkyi с ее сильно разрушенными задним и боковыми краями.
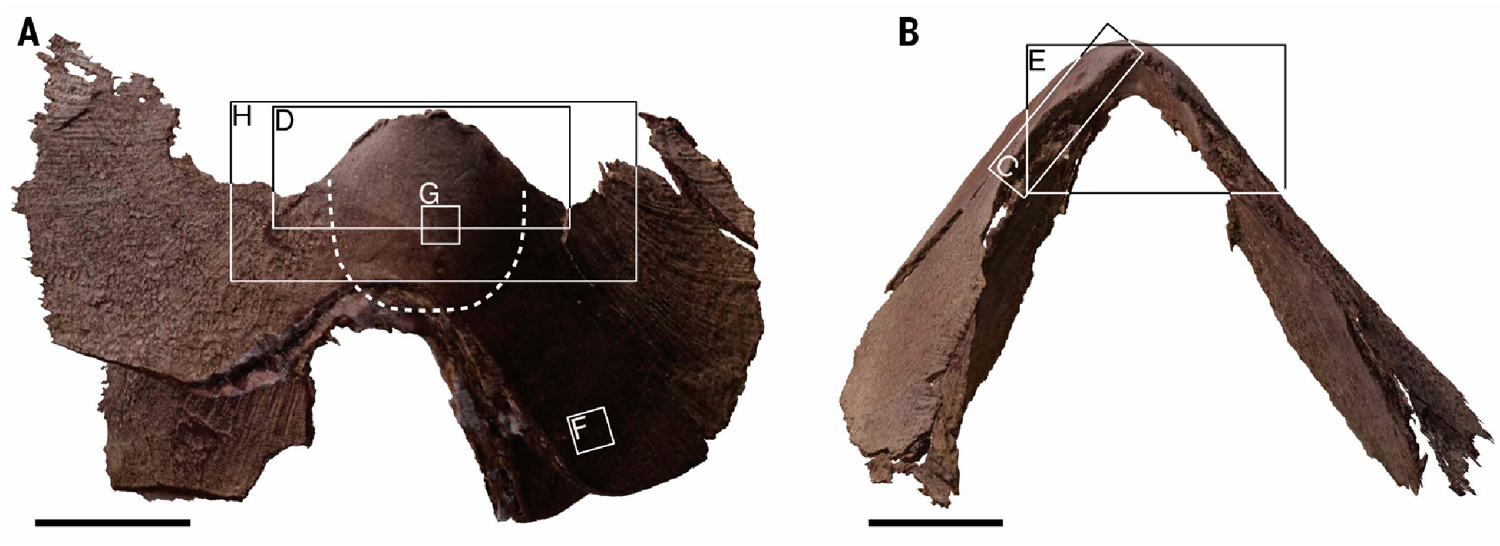
Полукруглое отверстие в задней части и многочисленные мелкие разрушения в краевых частях челюсти N. Jeletzkyi, свидетельствующие о неполной фоссилизации. Вполне вероятно, что челюсть подверглась довольно значительному разложению до того, как попала в условия, благоприятствующие фосфатизации. Снимки из обсуждаемой статьи.
Но мы помним, что именно форму челюсти авторы использовали для того, чтобы обосновать принадлежность нанаймотеутисов к цирратным осьминогам! Они использовали метод анализа главных координат (principal coordinates analysis), сравнивая с его помощью две челюсти Nanaimoteuthis с 19 челюстями современных осьминогов и вампиротеутиса. Сам выбор метода абсолютно логичен — вопросы к использованному материалу.
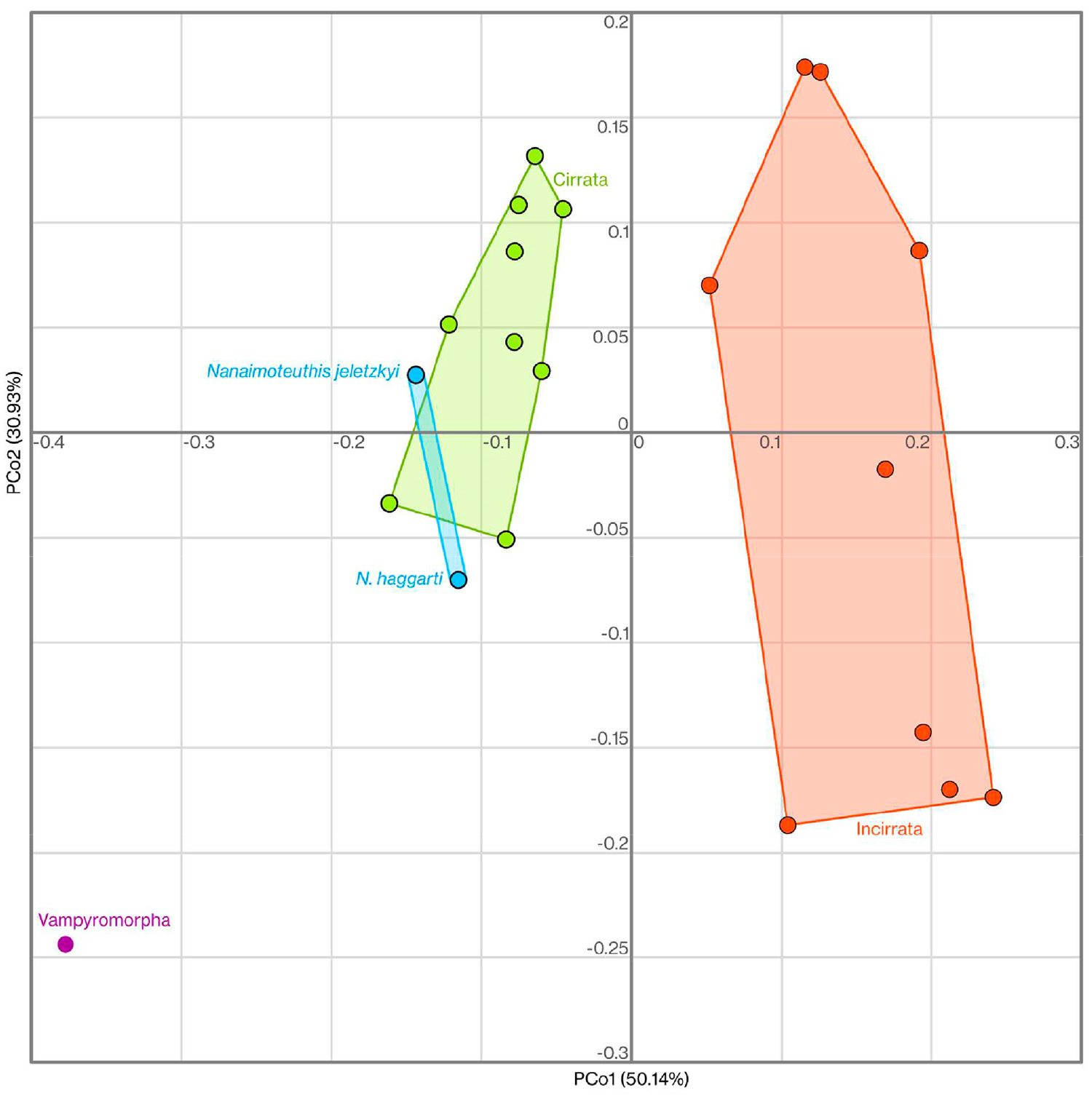
Диаграмма анализа главных координат, точки указывают положение видов в данном морфологическом пространстве. Подробнее см. дополнительные материалы к обсуждаемой статье
Судя по приведенной в дополнительных материалах к статье ссылке, авторы взяли данные по современным челюстям с веб-сайта “Tree of Life” (у меня ссылка не открывается, поэтому привожу «веб-архив»). Но там гораздо больше девятнадцати таксонов. Почему были выбраны именно эти 19 (их список приведен здесь)? Кроме того — почему авторы не взяли для сравнения ни одного ископаемого таксона челюстей? Ведь их описано немало.
И наконец самый главный вопрос: а в каком виде они сравнивали Nanaimoteuthis с современными прекрасно сохранившимися челюстями? В том, в каком он есть сейчас — с растворенными или истертыми элементами и посмертными деформациями? Но они очевидным образом искажают исходную форму челюсти. Или в том, каким он должен был бы быть изначально, с восстановленными не сохранившимися элементами (Fig. 1 в статье)?

Рисунок 1 из обсуждаемой статьи без челюсти современного кальмара. Обратите внимание на серые области, обозначающие не сохранившиеся части челюстей. Их размеры, да и в принципе их наличие, в статье никак не обоснованы, хотя они имеют принципиальное значение для определения таксономической принадлежности челюсти.
Но понимание изначального облика челюсти может быть разным: например, в обсуждаемой работе, судя по рисунку 1, авторы исходили из того, что внутренняя пластина челюсти (lateral wall на рисунке) была длинной, но ее задняя часть просто не сохранилась (показана серым). Исходя из чего были сделаны такие выводы, авторы не поясняют. А вот их предшественники (K. Tanabe et al., 2017. New records of coleoid cephalopod jaws from the Upper Cretaceous of Hokkaido, Japan, and their paleobiogeographic and evolutionary implications), считавшие Nanaimoteuthis челюстями вампироморф, полагали, что внутренняя пластина сохранилась почти полностью и изначально была короткой — именно этот признак характерен для челюстей современных вампиротеутисов, но не осьминогов. Почему авторы новой статьи с этим не согласились?

Нижние челюсти цирратных осьминогов 1 — Grimpoteuthis и 2 — Opistoteuthis, а также вампироморфы 3 — Vampyroteuthis. Видно, что они различаются длиной рукоятки (которая у вампироморфов очень короткая) и шириной крыльев капюшона. Однако задние части рукоятки и крыльев плохо сохраняются в ископаемом состоянии и для определения принадлежности челюстей крайне важна правильная реконструкция их исходной морфологии. Иллюстрация из статьи C. C. Lu, R. Ickeringill, 2002. Cephalopod beak identification and biomass estimation techniques: tools for dietary studies of southern Australian finfishes
Все эти вопросы без ответов приводят к тому, что ключевой момент статьи — принадлежность ископаемых находок челюстей именно к цирратным осьминогам — выглядит совершенно не доказанным. А ведь если обладатели челюстей были не цирратными осьминогами, а принадлежали к другой группе октобрахий, то и все приведенные в статье расчеты оказываются бессмысленным: другие октобрахии могли иметь совершенно иные пропорции тела и рук.

Stauroteuthis syrtensis, тот самый осьминог, чьи пропорции помогли исследователям получить для Nanaimoteuthis оценку длины в 18,6 метра, ловит рачков при помощи мембраны между руками. Тело Stauroteuthis адаптировано для его образа жизни, а не для того, чтобы быть высшим хищником. Рисунок из статьи M. A. Collins, R. Villanueva, 2006. Taxonomy, Ecology And Behaviour Of The Cirrate Octopods
Еще один момент, который очень смущает в данном исследовании — это прямой перенос пропорций современных осьминогов на их вымерших родственников без учета совершенно разного образа жизни и типа питания. Большинство современных цирратных осьминогов планктонофаги, свои длинные тонкие щупальца с натянутой между ними перепонкой они используют для ловли мелких рачков (К. Н. Несис, 2005. Головоногие: умные и стремительные). Им выгодно иметь длинные руки, чтобы охватывать натянутой между ними перепонкой наибольших объем воды с микроскопической добычей (на видео можно посмотреть, как это делает Cirroteuthis) . Но крупную добычу такими руками не удержишь. Для охоты на позвоночных им были бы нужны другие руки, в строении которых упор делался бы на силу, а не длину и растяжимость. И пропорции таких рук наверняка были бы иными.
И наконец, самое интересное: из отложений верхнего мела давно известны гигантские октобрахии с длиной мантии около двух метров! Причем это достоверные находки окаменелостей, а не гипотетические расчеты. Этих гигантов в настоящее время относят к роду Enchoteuthis, впервые описанному еще в 1968 году. Авторы новой статьи их даже упоминают, но только в контексте того, что эти древние октобрахии тоже были длиннотелыми. А ведь их странные гладиусы, формой и размерами похожие на лопату или весло, встречаются в меловых отложениях начиная с альба. То есть они возникают практически одновременно с появлением в палеонтологической летописи гигантских клювов.

Голотип Enchoteuthis (в центре) и самый крупный из извеcтных образцов (слева) в сравнении с человеком. Пропорции тела Enchoteuthis гипотетические, размеры и форма гладиусов (выделены белым) — реальные. Рисунок из Википедии
В настоящее время их, как и осьминогов, относят к отряду Octopoda, только не к цирратам или инцирратам, а к отдельному вымершему подотряду Teudopseina (D. Fuchs, 2020. Treatise Online no. 138: Part M, Chapter 23G: Systematic Descriptions: Octobrachia), среди представителей которого, по-видимому, скрываются и предки цирратных осьминогов. К сожалению, находок челюстей in situ, вместе с гладиусами, пока нет (либо на них не обращали внимания), но с учетом близкого родства теудопсеин и современных осьминогов, сходство их челюстей между собой было бы вполне ожидаемым. Так что Enchoteuthis или его ближайшие родственники выглядят вполне вероятными кандидатами на роль обладателей челюстей Nanaimoteuthis. Вряд ли они превосходили длиной современных гигантских кальмаров и вряд ли представляли какую-либо угрозу для крупных акул или морских рептилий, но в любом случае это были серьезные хищники, игравшие определенную роль в морских экосистемах позднего мела. И реально существовавшие, в отличие от девятнадцатиметровых осьминогов-«кракенов».
И в заключение стоит отметить, что начиная от заголовка и на протяжении всей статьи авторы постоянно подчеркивают, что их осьминоги будто бы «самые ранние» (“the earliest octopuses”). У читателя может создаться впечатление, что описываемые находки из сеномана принадлежат древнейшим осьминогам, и что осьминоги возникли примерно в это время — около 100 миллионов лет назад. Но это, мягко говоря, не так. Да, древнейшие находки отпечатков тел и гладиусов осьминогов происходят именно из сеноманских отложений — подробнее см. картинку дня Ископаемые осьминоги. Но один из родов этих сеноманских осьминогов, Styletoctopus, относится к современному семейству Octopodidae, его рудименты гладиуса («стилеты») имеют совершенно современную форму. Другой род сеноманских осьминогов, Keuppia, относится к вымершему семейству Palaeoctopodidae, его гладиус уже разделился на две части, но еще не редуцировался до состояния стилетов. Разнообразие сеноманских осьминогов и наличие среди них форм современного типа говорит о том, что к этому времени они уже прошли очень большой эволюционный путь. А по данным молекулярных часов, два современных рода — Octopus и Enteroctopus — разошлись еще 150 миллионов лет назад — в поздней юре (B. Kröger et al., 2011. Cephalopod origin and evolution: A congruent picture emerging from fossils, development and molecules: Extant cephalopods are younger than previously realised and were under major selection to become agile, shell-less predators). А ведь это инцирратные осьминоги, которые в эволюционном плане явно моложе цирратных. Так что настоящие самые древние осьминоги жили как минимум уже в средней юре, а может быть и раньше.
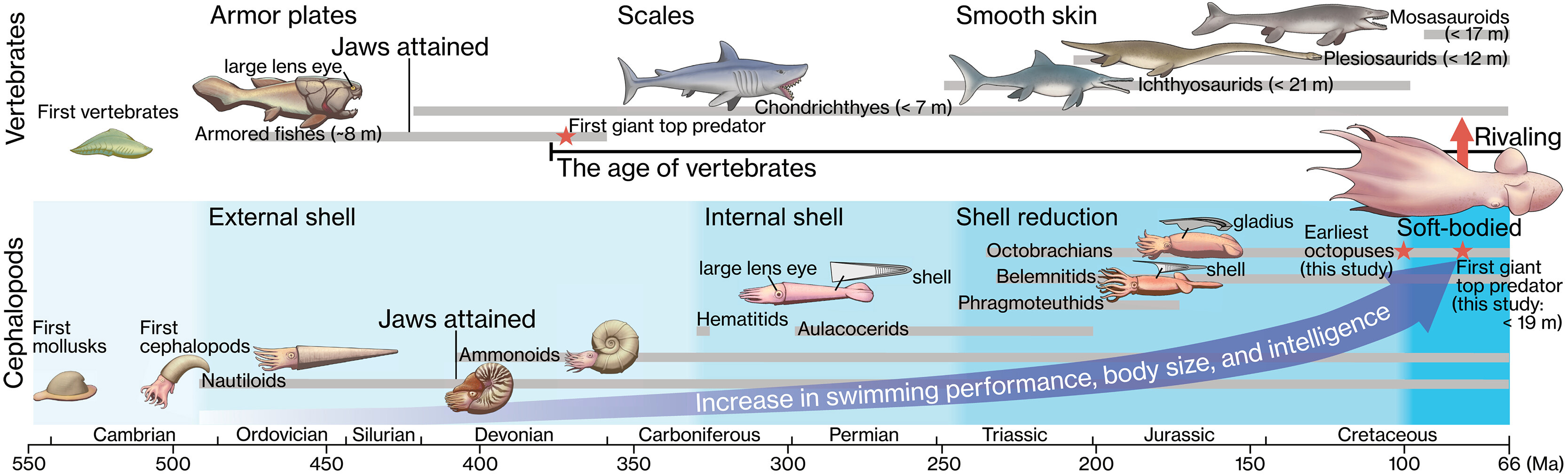
Одна из иллюстраций к статье, глядя на которую можно подумать, будто осьминоги возникли в середине мелового периода
Александр Мироненко
Последние новости




















{kind=link}
В глубинах древнего моря сто миллионов лет назад гигантский осьминог при помощи биолюминесценции охотится на хищную морскую рептилию мозазавра. Рисунок Julius Csotonyi