Территориальная агрессия у птиц-дуплогнездников формируется разными путями

Авторы статьи, опубликованной в журнале Nature Ecology & Evolution, изучали конвергентную эволюцию на примере территориальной агрессии к конспецификам, свойственной многим птицам, облигатно гнездящимся в дуплах. Ученые выбрали по паре видов из пяти семейств — в каждой паре один вид был облигатным дуплогнездником, а второй — нет. Птиц исследовали на трех уровнях — поведенческом, биохимическом и молекулярном (экспрессия генов в мозге). Оказалось, что тестостерон — традиционный «ответственный» за формирование агрессии — не ассоциирован напрямую со степенью территориальной агрессивности птиц (по крайней мере, у десяти исследованных видов). Несмотря на то что гены, ассоциированные с территориальным поведением, в основном разные у разных видов, авторы обнаружили общие закономерности в экспрессии небольшого набора генов, активных в мозге, — в том числе сети генов, схожей у нескольких видов-дуплогнездников. Таким образом, показано, что конвергенция в данном случае обусловлена как сочетанием схожей эволюции сходных генов, так и независимым возникновением молекулярных механизмов, основанных на негомологичных генах.
Конвергентная эволюция — возникновение у неродственных организмов схожих фенотипических признаков, обусловленное жизнью в схожих условиях. Конвергенция может проявляться как в морфологии (то есть во внешнем виде или особенностях строения), так и менее очевидно — например, в поведении. Ученых интересуют не только отдельные случаи конвергенции, но и молекулярно-генетические процессы, которые ее сопровождают. Ведь, с одной стороны, конвергенция может быть результатом эволюции одних и тех же молекулярных и биохимических механизмов через гомологичные гены, подвергшиеся схожему влиянию отбора. С другой стороны, конвергенция может оказаться удачным сочетанием действия принципиально разных генов или химических веществ. В разных исследованиях показано, что оба варианта встречаются в природе. Поэтому, изучая виды со сходными фенотипическими признаками, без дополнительных исследований невозможно достоверно определить, какой эволюционный «маршрут» они выбрали. Важно также учитывать степень эволюционного родства между изучаемыми видами, чего обычно не делают.

Рис. 2. Представители семейства Древесницевые: слева — лимонный певун (Protonotaria citrea), облигатный дуплогнездник: справа — желтая древесница (Setophaga petechia), которая обычно строит гнезда на ветках деревьев. Фото с сайта phys.org
Авторы недавней статьи в журнале Nature Ecology & Evolution постарались избавиться от этих методологических недостатков и учесть одновременно поведенческие, гормональные и нейрогеномные (см. Neurogenomics) особенности. В качестве объектов исследования они выбрали по паре видов птиц из 5 семейств отряда Воробьинообразные (Passeriformes) — Ласточковые (древесная американская ласточка (Tachycineta bicolor) и деревенская ласточка (Hirundo rustica)), Древесницевые (лимонный певун (Protonotaria citrea) и желтая древесница (Setophaga petechia)), Воробьиные (полевой воробей (Passer montanus) и домовый воробей (Passer domesticus)), Дроздовые (восточная сиалия (Sialia sialis) и странствующий дрозд (Turdus migratorius)) и Крапивниковые (домовый крапивник (Troglodytes aedon) и каролинский кустарниковый крапивник (Thryothorus ludovicianus)). При выборе «подопытных» ученые исходили из того, что, во-первых, виды в каждой паре должны быть эволюционно близки друг к другу, а во-вторых, один из них должен строить гнезда исключительно в дуплах, а второй вид должен иметь более гибкую стратегию гнездования — либо пользоваться дуплами при случае, либо вообще не гнездиться в них (в перечислении выше всюду первым указан вид, облигатно гнездящийся в дуплах).
Для птиц-дуплогнездников, облигатно гнездящихся в дуплах деревьев (или других укромных полостях), подходящие места для гнезд — очень ценный и дефицитный ресурс, за который порой разгорается нешуточная конкуренция как между конспецификами, так и между представителями разных видов (см., например, новость Острота конкуренции за гнезда между синицами и мухоловками зависит от климатических условий, «Элементы», 14.02.2019 и картинку дня Узурпаторы гнезд). Видимо, из-за этого птицам-дуплогнездникам свойственен довольно высокий уровень территориальной агрессии. У позвоночных, предположительно, в формировании агрессии задействованы гормон тестостерон, а также его метаболит 17-бета-эстрадиол, активирующие стероидные рецепторы в мозгу. Но помимо них есть и другие химические источники агрессии: в мозгу экспрессируется большое число генов, белковые продукты которых (а также их концентрация) способны влиять на поведение животного, в том числе — и на его уровень агрессии. Так, например, на поведение могут влиять профили экспрессии G-белков, дофамина, серотонина и глутамата, а также другие гены, часто встречающиеся даже у неблизкородственных видов. При изучении таких генов необходимо делать поправку на вариации в экспрессии генов как между видами, так и между самцами и самками в пределах одного вида (авторы обсуждаемого исследования это учитывали).
Поведенческий уровень
Чтобы количественно измерить территориальную агрессию птиц, ученые помещали на скворечник (а для недуплогнездников — около мест, где они чаще всего проводили время) чучело птицы того же вида и беспроводную колонку, проигрывающую пение птицы того же вида. Через 30 секунд после проигрывания аудиозаписи ученые в течение 5 минут следили за ситуацией и отмечали агрессивное поведение в отношении чучела со стороны исследуемой птицы, особенно фокусируясь на физическом контакте. Полученные результаты соотносили со стратегией гнездования, полом птицы и полом чучела в линейной модели и корректировали в соответствии с филогенетическим родством изучаемых птиц.
Оказалось, что если объединить результаты наблюдений по всем семействам, то дуплогнездники значительно более агрессивно относятся к чучелу-конспецифику, чем птицы, не гнездящиеся в дуплах. Причем у облигатных дуплогнездников в такой агрессии замечены и самки, и самцы, а у факультативных — в основном только самцы (рис. 3, b). Птицы, гнездящиеся открытым способом, практически не проявляют территориальной агрессии к конспецификам. У облигатных дуплогнездников самцы в среднем немного более агрессивны, причем авторы отмечают, что они чаще атаковали чучела самцов, в то время как самкам пол чучела был не так важен. По отдельным семействам ситуация менее однозначная — см. рис. 3 и подпись к нему.
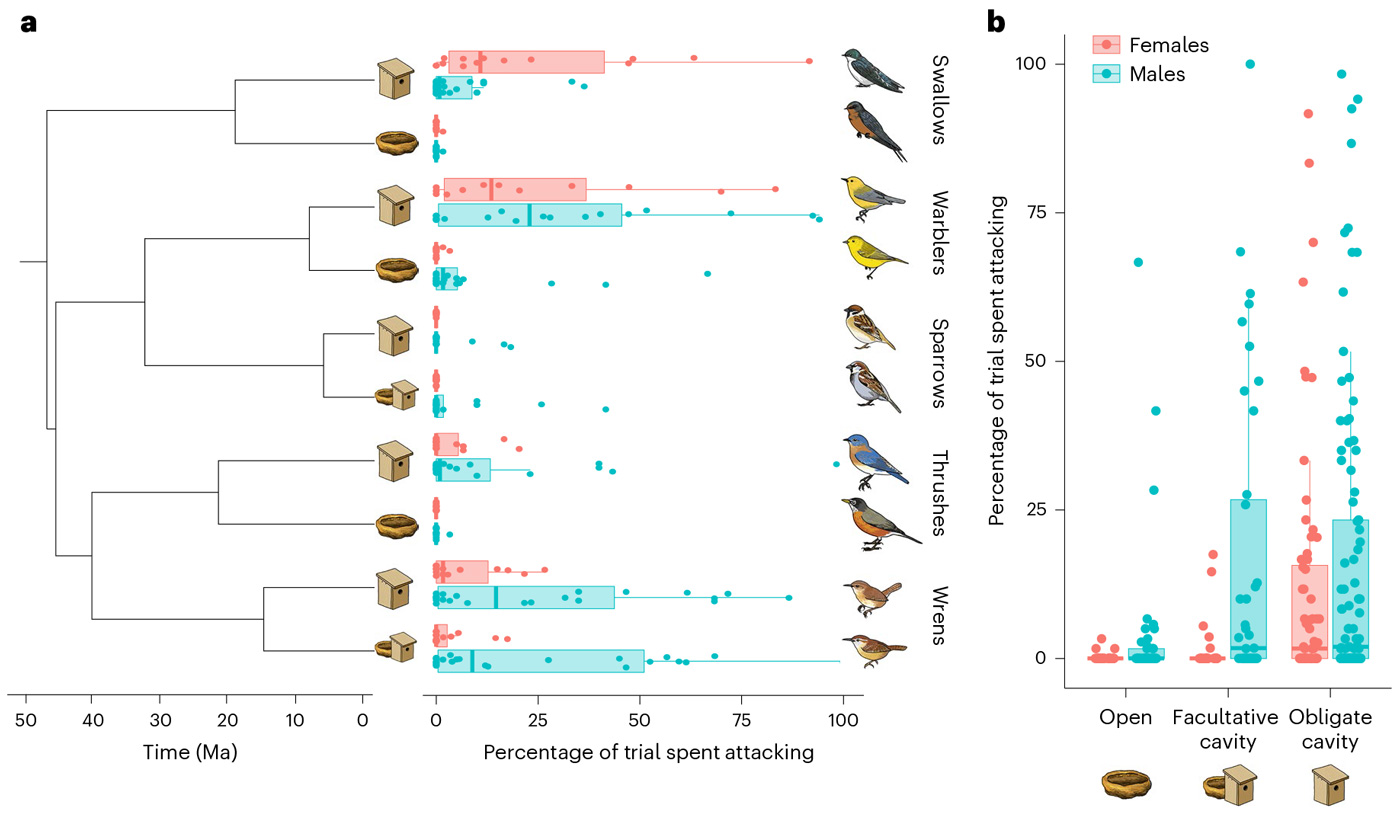
Рис. 3. Результаты наблюдений и оценка агрессивности исследуемых видов. a — филогенетическое древо и разбивка данных по виду и полу (красным — самки, голубым — самцы). Изученные семейства разошлись от общего предка около 44 миллионов лет назад, пары видов в пределах одного семейства разошлись от 9 до 20 миллионов лет назад. Видно, что в парах, в которых один из видов отчетливо проявляет территориальную агрессию к конспецификам, — то есть ласточек, древесниц и дроздов — это делают именно дуплогнездники; исключениями здесь служат миролюбивые воробьи (оба вида практически не проявляют агрессии) и воинственные крапивники (оба исследованных вида оказались весьма агрессивными). Виды птиц по семействам сверху вниз: Ласточковые — древесная американская ласточка (Tachycineta bicolor) и деревенская ласточка (Hirundo rustica), Древесницевые — лимонный певун (Protonotaria citrea) и желтая древесница (Setophaga petechia), Воробьиные — полевой воробей (Passer montanus) и домовый воробей (Passer domesticus), Дроздовые — восточная сиалия (Sialia sialis) и странствующий дрозд (Turdus migratorius), Крапивниковые (домовый крапивник (Troglodytes aedon) и каролинский кустарниковый крапивник (Thryothorus ludovicianus). b — Агрессия к чучелу своего вида, разделенная по стратегии гнездования и полу. Самки облигатных дуплогнездников оказались значительно агрессивнее самок видов, предпочитающих другие стратегии гнездования. При этом как облигатные, так и факультативные самцы-дуплогнездники агрессивнее самцов видов с открытым гнездованием. Рисунок из обсуждаемой статьи
Биохимический уровень
Следующим шагом стало измерение тестостерона у птиц — чтобы выяснить, действительно ли он влияет на уровень территориальной агрессии. Из предыдущих исследований, например, было известно, что у самок древесных американских ласточек уровень тестостерона коррелирует с территориальной агрессией, в то время как у самцов такой связи не прослеживается. Про другие виды не было известно, существует ли связь между тестостероном и агрессивным поведением. Поэтому авторы решили разобраться и в этом. Они установили ловушки поблизости от гнезд. Часть отловленных птиц они не провоцировали перед тем, как взять образцы крови и мозга, у части провоцировали агрессивное поведение, тут же умерщвляли их и собирали образцы, третью же группу провоцировали на территориальную агрессию и собирали образцы через несколько дней после этого.
При объединении результатов анализа образцов всех видов оказалось, что не существует значимой разницы между уровнем тестостерона и агрессивным поведением как у самцов, так и у самок. У видов, которые были существенно представлены в исследовании (восточная сиалия, домовой крапивник, каролинский кустарниковый крапивник, лимонный певун, желтая древесница) по отдельности также не оказалось существенной связи между тестостероном и агрессией. Однако, как и ожидалось, у самцов обнаруживался значительно более высокий уровень тестостерона.
Молекулярный уровень
Если агрессия у птиц закладывается не на биохимическом уровне, это означает, что причину надо искать в более глубоких и тонких процессах, например в разнице в экспрессии генов в мозге. Такая связь может быть не всегда очевидна, поскольку многие белки выполняют несколько функций сразу, а точная функция и механизм работы некоторых генов до сих пор не известны — известно лишь, что они экспрессируются в определенных органах или ассоциированы с определенным процессом.
Исследователи измерили количество матричной РНК 10 672 различных генов, которые экспрессируются у всех изученных видов в вентромедиальной части префронтальной коры (ventromedial prefrontal cortex), регулирующей, в том числе, агрессию. Они обнаружили, что часто в пределах пары сравниваемых видов одного семейства весомая доля генов (в среднем — 1390 генов, или 13% от общего количества исследованных генов) отличается по уровню экспрессии у облигатного дуплогнездника и у птицы с факультативным/открытым гнездованием. Однако при сравнении нескольких семейств между собой выяснилось, что часто гены, по-разному экспрессирующиеся в парах видов, не совпадали. Общими для всех пяти семейств оказались только 11 генов (0,1%) из всего изученного разнообразия. С одной стороны, это показывает, что большая часть молекулярных механизмов, которые приводят к формированию территориальной агрессии, отличается у изученных видов, а с другой стороны, 11 генов — намного больше, чем ожидали исследователи (по их расчетам, в рандомизированной модели нашелся бы всего один общий ген). Кроме того, обнаруженные 11 генов образуют белки, для которых ранее была установлена связь с регуляцией поведения — EMC3, NPTX1 и RGS19 связаны с рецепцией нейромедиаторов, ATP6V0E — с активностью АТФазы, ARRDC4, CTNND1, NPTX1, WIPF2 — с развитием болезни Альцгеймера, RGS19 — с формированием зависимости, AFP1L1 и PQBP1 — с нейродегенеративными заболеваниями. У этих генов также присутствовали общие регуляторные мотивы (короткие фрагменты ДНК, которые узнают факторы транскрипции и которые определяют, в каких условиях запускается или отключается экспрессия гена). Эти мотивы — E2F, ассоциированный с нейрогенезом, и ETS-1, ассоциированный с поддержанием жизненных процессов клетки.

Рис. 4. a — число одинаково экспрессируемых генов, общих между семействами (то есть между тепловыми картами) для самцов (синим), самок (розовым) и обоих полов вместе (фиолетовым). Сплошные линии обозначают более высокую экспрессию у обязательных дуплогнездников, пунктирные — более низкую. У всех пяти семейств наблюдается разница в экспрессии у 11 общих генов. b — пояснение к рисунку c: в левой верхней части тепловой карты показаны гены, пониженно экспрессирующиеся у первого семейства и повышенно у второго, в правой верхней — повышено у обоих семейств, в левой нижней — пониженная экспрессия у обоих семейств, в правой нижней — повышенная у первого, пониженная у второго. c — в каждом пикселе тепловой карты — наборы из пар около 100 генов. Чем теплее цвет пикселя, тем больше пар генов экспрессируются скоординировано между двумя семействами. Рисунок из обсуждаемой статьи
Авторы также составили линейную модель, задачей которой было выяснить, ассоциированы те или иные гены со стратегией гнездования, агрессией, полом и парами признаков «пол + стратегия» и «агрессия + стратегия». В данную модель также внесли корректировки, целью которых было избежать ложно-положительного результата при сравнении эволюционно близких семейств. Из более чем 10 000 изученных генов 234 оказались ассоциированы с облигатным гнездованием в дуплах, 79 — с агрессией, из них 5 генов были ассоциированы с обоими факторами. Еще 62 гена были ассоциированы с совместным действием этих факторов, 76 — с полом и облигатной дуплогнездностью, причем примерно одинаковое количество генов было больше ассоциировано с каждым из полов. При этом пол влияет на экспрессию наибольшего числа генов — 510, большинство их этих генов активнее экспрессируется у самцов и многие из них отвечают за клеточный метаболизм. Среди 234 генов, ассоциированных с дуплогнездностью, 168 встречаются в как минимум трех семействах и отличия в их экспрессии от не-дуплогнездников выражены во всех семействах в одном направлении. Во всех пяти семействах в одинаковом направлении менялась экспрессия 50 генов, причем гены, ответственные за тестостерон и стероидные гормоны не ассоциированы со стратегией гнездования, в отличие от, например, генов, которые (сами или их гомологи) ассоциированы с АТФ, функцией митохондрий или психиатрическим риском.
Изучая молекулярные основы формирования поведения, важно помнить, что часто на поведение гены влияют не в одиночку, а образуя скоординированную сеть (gene co-expression network), причем гены, входящие в такую сеть тоже могут меняться в ходе эволюционных процессов. Поэтому авторы исследовали коэкспрессию групп генов для всех 10 видов, чтобы выяснить, присутствуют ли в изученном материале сети генов и влияют ли они на поведение птиц. Изначально они обнаружили 93 сети, однако после составления линейной модели, выявляющей связь этих сетей с полом, агрессией, стретагией гнездования и взаимодействиями этих факторов, и поправки на филогенетические связи, авторы обнаружили всего две сети (red и tan4), ассоциированные с этими факторами, существование которых нельзя объяснить случайными процессами генетического дрифта. Сеть tan4 ассоциирована со стратегией гнездования и содержащая ряд генов, связанных с работой митохондрий, метаболизмом, когнитивными функциями и ответами на стресс (рис. 5).

Рис. 5. Tan4 — сеть генов, ассоциированная с облигатной дуплогнездностью. b — тепловая карта генов, включенных в сеть; отдельные колонки — индивидуальные особи, сгруппированные по полу и стратегии гнездования, цвет — уровень экспрессии одного из 44 генов сети. c — 44 гена, включенные в сеть и ассоциации между ними
При этом у самок были обнаружены две специфичные только для них сети генов, ассоциированные с агрессией, — pink4 и sienna4, включающие гены, связанные с дофаминовыми и глутаматовыми рецепторами. Наличие таких сетей показывает, что агрессивное территориальное поведение регулируется несколькими механизмами, которые могут отличаться как между отдельными видами, так и между полами в пределах одного вида.
Рис. 6. Сети генов pink4 (c) и sienna4 (d). Рисунок из обсуждаемой статьи
Вероятно, разница в экспрессии генов между полами отчасти объясняется тем, что у птиц самцы — гомогаметный пол. В отличие от, например, млекопитающих, у которых самцы обычно несут XY-хромосомы, а самки — XX-хромосомы, у птиц самцы являются носителями ZZ-хромосом, а самки — ZW-хромосом. Поэтому гены, находящиеся на второй Z-хромосоме самцов, могут экспрессироваться значительно больше, чем у самок, особенно если у самцов отсутствуют механизмы дозовой компенсации, предотвращающие данный процесс. Действительно, например, многие гены сети red находятся на Z-хромосоме. Другие возможные объяснения этого процесса — более сильное влияние на самцов полового отбора, эволюционные ограничения на снижение уровня генетической экспрессии. Все эти данные указывают на то, что механизмы развития территориальной агрессии хотя бы частично отличаются между самцами и самками.
Обсуждаемая работа — подробный анализ формирования конвергенции в поведенческих процессах. Она показала, что территориальная агрессия формируется на более глубоком уровне, чем просто биохимический. Почти у всех изученных пар видов облигатные дуплогнездники агрессивнее реагировали на чучело своего вида (причем самки атакуют чучела обоих полов, а самцы — только своего). В изученных видах тестостерон, судя по всему, не влияет на уровень агрессии (в противовес распространенной гипотезе о биохимических механизмах ее формирования), а является лишь одним из факторов, определяющих территориальное поведение.
Источник: Sara E. Lipshutz, Mark S. Hibbins, Alexandra B. Bentz, Aaron M. Buechlein, Tara A. Empson, Elizabeth M. George, Mark E. Hauber, Douglas B. Rusch, Wendy M. Schelsky, Quinn K. Thomas, Samuel J. Torneo, Abbigail M. Turner, Sarah E. Wolf, Mary J. Woodruff, Matthew W. Hahn & Kimberly A. Rosvall. Repeated behavioural evolution is associated with convergence of gene expression in cavity-nesting songbirds // Nature Ecology & Evolution. 2025. DOI: 10.1038/s41559-025-02675-x.
Владимир Вержбицкий
-
> Таким образом, показано, что конвергенция в данном случае обусловлена как сочетанием схожей эволюции сходных генов, так и независимым возникновением молекулярных механизмов, основанных на негомологичных генах.
This sounds very much like AI generated bullshit.-
))
Или электронный перевод.
В любом случае: тут Вы подметили местечко, в котором наблюдается присутствие полного отсутствия эмпатии к читателю, к его трудностям понимания. Как один знакомый юрист сказал на пожелание привести зубодробительный текст к состоянию, когда сложносочиненные-подчиненные-перечиненные предложения будут разрываться точкой хотя бы единожды за абзац: "зачем? Юристы и так поймут!"
-


То есть агрессия против чучелки рядом с домом волне может быть вызвана не желанием защитить дом от конкурента-собрата, а, например, праведным гневом по адресу натуралистов-исследователей, которые не дают покоя семье, включая громко музыку и подбрасывая в наш двор этот хлам - дурацкие здоровенные чучела. "Уберите нафиг!"
Я как птица сомневаюсь, что спутаю статую командора перед домом, даже обклеенную перьями для правдоподобия, от соседа, с гитарой поющего под окном романсы, масляно поглядывая на мою жену...
Но если б знал, что эти юннаты еще и ловить нас собираются и убивать ради пробы тканей мозга (да еще и перед убийством будут мучать-злить) - всеми силами стремился бы выклевать им глаза.
"Часть отловленных птиц они не провоцировали перед тем, как взять образцы крови и мозга, у части провоцировали агрессивное поведение, тут же умерщвляли их и собирали образцы"
Кто тут про ужас птичий говорил?
-
Ну да.
Дразнили, провоцировали, а потом убивали и брали образцы мозга.
И ещё получали от этого удовольствие.
Трижды получали удовольствие.
Первый раз когда дразнили, зная что потом убьют.
Второй раз когда убивали.
И третий раз получали удовольствие вскрывая череп и беря образцы мозга.
Это типичное поведение многоклеточных гетеротрофов.
Была заблуждением попытка поправить эту ветвь эволюции убрав с арены самую кусучую их часть 66 млн лет назад.
Народились новые.
Более отвратительные, умные и безжалостные.
С многочисленными многоклеточными гетеротрофами нужно кончать раз и навсегда.
Комета уже летит.
"
одна из птиц-дупогнездников, изученных в обсуждаемой работе"
Последние новости













Рис. 1. Слева — древесная американская ласточка (Tachycineta bicolor) — одна из птиц-дуплогнездников, изученных в обсуждаемой работе. Справа — деревенская ласточка (Hirundo rustica), которая предпочитает строить гнезда вне дупел или других полостей. Фото с сайтов en.wikipedia.org и birdsoftheworld.org